Influence of Betaxolol on the Methamphetamine Dependence in Mice
Article information
Abstract
Objective
The noradrenaline system is involved in the reward effects of various kinds of abused drugs. Betaxolol (BTX) is a highly selective β1-antagonist. In the present study, we evaluated the effect of BTX on methamphetamine (MAP)-induced conditioned place preference (CPP) and hyperactivity in mice.
Methods
The mice (n=72) were treated with MAP or saline every other day for a total of 6 days (from day 3 to day 8; 3-times MAP and 3-times saline). Each mouse was given saline (1 mL/kg) or MAP (1 mg/kg, s.c.) or BTX (5 mg/kg, i.p.) or MAP with BTX (5 mg/kg, i.p.) 30 min prior to the administration of MAP (1 mg/kg, s.c.) every other day and paired with for 1 h (three-drug and three-saline sessions). We then compared the CPP score between the two groups. After the extinction of CPP, the mice were given BTX (5 mg/kg, i.p.) or saline (1 mL/kg) 24 h prior to a priming injection of MAP, and were then immediately tested to see whether the place preference was reinstated.
Results
The repeated administration of BTX 30 min prior to the exposure to MAP significantly reduced the development of MAP-induced CPP. When BTX was administered 24 h prior to the CPP-testing session on day 9, it also significantly attenuated the CPP, but did not result in any change of locomotor activity. In the drug-priming reinstatement study, the extinguished CPP was reinstated by a MAP (0.125 mg/kg, s.c.) injection and this was significantly attenuated by BTX.
Conclusion
These findings suggest that BTX has a therapeutic and preventive effect on the development, expression, and drug-priming reinstatement of MAP-induced CPP.
INTRODUCTION
Drug addiction has important biological, psychological, and social causes and consequences, and it has been recognized as a psychiatric disease that involves the development of complex behaviors such as tolerance, dependence, and cravings regarding drugs that are characteristic of an addictive state. Methamphetamine (MAP) has strong psychostimulant effects and a severe addictive property, and its abuse is a major health problem; moreover, MAP addiction has no corresponding detoxification agent yet.1
Although the inhibition of dopamine (DA) uptake and the subsequent stimulation of DA receptors are established mechanisms that mediate the abuse-related effects of psychostimulants, existing evidence shows that noradrenaline (NE) uptake and NE-receptor mechanisms can also have important modulating roles regarding the behavioral effects of psychostimulants in laboratory animals. In pigeons, rats, and monkeys that have been trained to discriminate relatively low doses of cocaine from a vehicle, NE-transport inhibitors partially mimic the discriminative stimulus effects of cocaine,23456 and when combined with cocaine or the selective DA-transport inhibitor GBR 12909, the NE-transport inhibitors enhanced these discriminative stimulus effects.267 Furthermore, the discriminative stimulus effects of cocaine and the cocaine-like effects of NE-transport inhibitors such as talsupram and tomoxetine can be attenuated by the alpha-adrenoceptor antagonist prazosin.46 Previous studies showed that those animal subjects that received the beta-adrenoceptor antagonist propranolol following a preference test showed no preference for the cocaine-paired floor during a subsequent test, whereas vehicle-treated rats continued to express a preference for the cocaine-paired floor.8 These results suggest that the propranolol induced an impairment with respect to drug-seeking behavior.
Since propanolol, which is both a β1- and β2-adrenergic receptor antagonist, has a binding capability to 5-HT (serotonin) receptors,9 and atenolol, a selective β1 antagonist, enters the brain in very limited amounts,10 these drugs may affect MAP-induced CPP and hyperactivity via a blocking of the peripheral β1-adrenergic receptors.11 To avoid the effect of the β1-adrenergic receptor antagonist via the peripheral receptors, we used betaxolol (BTX) in our study since BTX easily crosses the blood-brain barrier.12 BTX is a highly selective β1 antagonist with no partial agonist (intrinsic sympathomimetic) activity and weak membrane-stabilizing (local anaesthetic) activity.13 Place conditioning is commonly used to measure the rewarding or incentive properties of drugs, especially regarding psychological dependence. The conditioned place preference (CPP) model can therefore be used as an effective tool to investigate the underlying mechanisms of a drug-induced reinstatement of drug-seeking behavior after extinction,14 and can partly evaluate the associated learning ability. In the present study, we investigated the effect of BTX on MAP addiction with a sample of MAP-induced mice, whereby a CPP development, expression, and reinstatement model was used to elucidate the effect of BTX on drug addiction and relapse.
METHODS
Animals
Male ICR mice (n=72; aged from 4 weeks to 5 weeks; Samtako Bio, Korea) were used in the CPP test. All of the animals were maintained under a constant temperature of 22±2℃, and were kept on a 12 h light/12 h dark cycle. Food (standard laboratory chow) and water were available ad libitum. All possible efforts were made to minimize animal suffering and the number of animals that were used, in accordance with the Chonbuk National University Medical School's "Guidelines for Animal Experiments."
Apparatus
The dimensions of the plexiglass CPP boxes measured 30×15×30 cm–width (w), length (l), and height (h), respectively–and the insertion of a removable clipboard divided the boxes into two equal-sized compartments (15×15×30 cm). One compartment had black and white striped walls and a black grid floor (chamber A), and the other compartment had black walls and a smooth floor (chamber B). In our experimental condition, the mice showed a significant spontaneous preference for chamber A, and we therefore used a biased method, whereby chamber B served as the MAP-paired compartment. The entire experimental process was documented using a video tracking system (Smart-CS, Panlab, Spain).
Procedures
Effect of BTX on development of MAP-induced CPP
On the first day, each mouse habituated by freely exploring the two compartments for 20 min without any treatment. On the second day, we tested the natural preferences of the mice. The animal subjects were allowed to freely explore both compartments for 20 min, and the compartment that was occupied for a lesser time was appointed as the drug-paired side (compartment B). The mice that received MAP were subjected to the least-preferred side. The biased method15 that we used meant that the MAP-paired compartment was initially the non-preferred side of the apparatus, as was measured during the pre-test.16
Between day 3 and day 8, the mice were divided into four groups; each mouse was given saline (1 mL/kg) or MAP (1 mg/kg, s.c.) or BTX (5 mg/kg, i.p.) or MAP with BTX (5 mg/kg, i.p.) 30 min prior to the administration of MAP (1 mg/kg, s.c.) on day 3, 5, 7 in compartment B, and all mouse was given saline (1 mL/kg) on day 4, 6, 8 in compartment A. Methamphetamine (MAP) was purchased from Dainippon Sumitomo Pharma (Osaka, Japan). During the 6 days of the experiment, the locomotor activity of each group was also monitored. The behavior of the mice was recorded by a video camera and analyzed by Smart-CS system set (Panlab, Barcelona, Spain). On the test day (day 9), the animals were allowed free access to both compartments. The time spent in each compartment was recorded for 20 min.
Effect of BTX on established MAP-induced CPP
To assess the blocking effect of BTX on MAP-induced CPP, each animal was conditioned with MAP (1 mg/kg, s.c.) or saline (1 mL/kg) without the pretreatment of BTX that was previously described. The mice were administered BTX (5 mg/kg, i.p.) or saline next day (24 h after) of the final conditioning.17 CPP were then tested 2 day (48 h) after the final conditioning.18
Effect of BTX on drug-priming reinstatement of MAP-induced CPP in mice
Following the development of CPP, all of the mice were retested without any treatment every 2 days to examine the maintenance of CPP. Two days after determining the extinction of CPP,16 they were given BTX (5 mg/kg, i.p.) or saline. After 24 h, each animal was injected with a priming dose of MAP (0.125 mg/kg, s.c.) or saline and were immediately tested to see whether the CPP was reinstated.
Statistical analysis
Data are expressed as mean±SEM. The CPP score was calculated as the difference between the times spent in chamber B during the pre- and post-conditioning phases. The significance of the difference between the groups was determined by a one-way ANOVA, followed by either the Manny-Whitney U-test (between 2 groups) or the Kruskal-Wallis test (among 4 groups), using GraphPad Prizm 3.0 (GraphPad Software, U.S.) software, which was commercially available at the time of the experiment. Probability values <0.05 were considered significant.
RESULTS
In this experiment, we assessed the effect of BTX on the development of MAP-induced CPP in mice. The 3 pairings of MAP (1 mg/kg, i.p.) with the naturally non-preferred chamber induced significant conditioning for the MAP-paired chamber. There was a significant increase of the time spent in the MAP-paired chamber with respect to the control group that was treated with saline instead of MAP (p<0.05; Kruskal-Wallis test). As shown in Figure 1, the pretreatment of BTX (5 mg/kg, i.p.) significantly blocked MAP (1 mg/kg, s.c.)-induced CPP (p<0.01; Kruskal-Wallis test). On its own, the BTX was devoid of an effect regarding place preference at the dose employed in this study. One day after the 3 pairings with MAP (1 mg/kg, s.c.), the time spent in the MAP-paired compartment was increased by approximately 18%, meaning that MAP-induced CPP was developed and maintained.
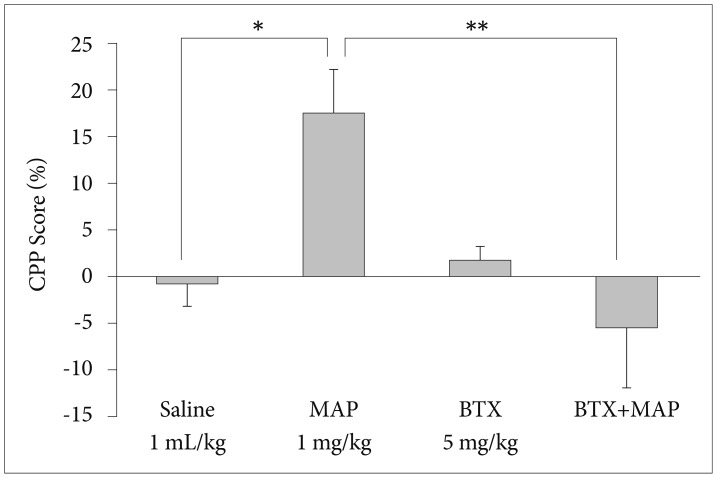
Effect of betaxolol (BTX, 5 mg/kg, i.p.) on development of methamphetamine (MAP, 1 mg/kg, s.c.) induced CPP in mice. Data are presented as mean±standard error of the mean (each group N=6). *p<0.05, **p<0.01. MAP+BTX: BTX (5 mg/kg, i.p.) injection 30 min prior to the MAP injection. CPP: conditioned place preference.
The repeated treatment of MAP gradually increased the locomotor activity of the mice, which was not affected by the BTX (Figure 2). Figure 2 shows that the repeated treatment of MAP or BTX+MAP displayed a significantly enhanced locomotion in response to the same dose of MAP on day 3 and day 5 compared with day 1, indicating a behavioral sensitization. The BTX-or-saline-treated group did not show any sensitization across the test sessions (p>0.05; one-way ANOVA). As indicated in Figure 3, the single administration of BTX (5 mg/kg, i.p.) 24 h prior to the CPP test significantly attenuated the maintenance of MAP-induced CPP in the mice (p<0.05; Kruskal-Wallis test).
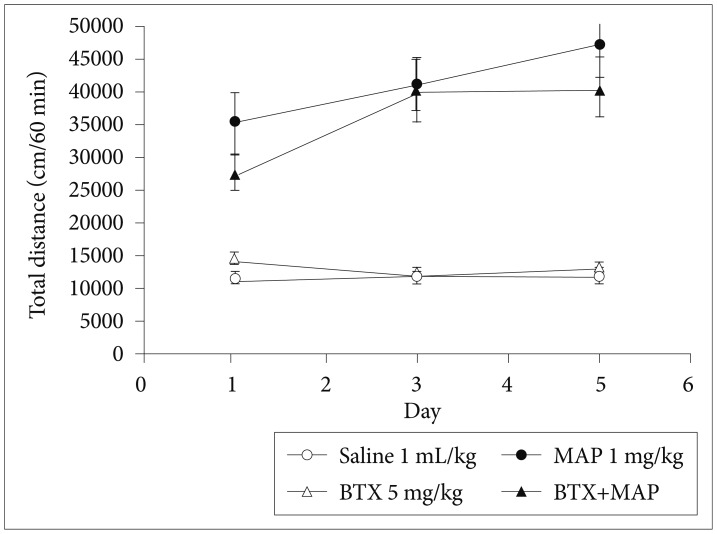
Effect of betaxolol (BTX, 5 mg/kg, i.p.) on the methamphetamine-induced locomotor activity in mice. Data are presented as mean±standard error of the mean (each group N=6).
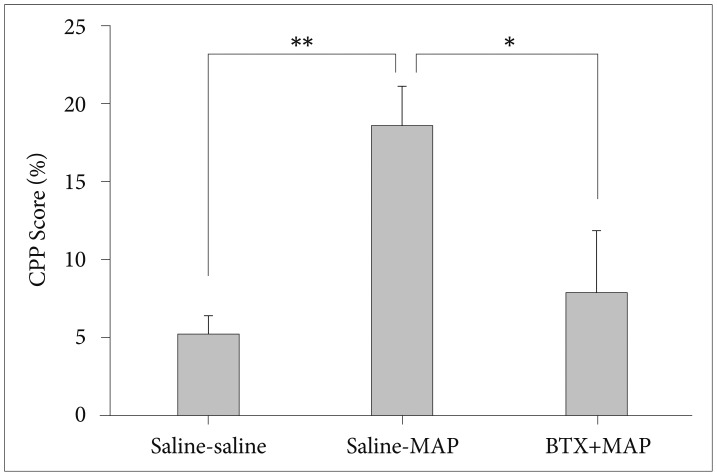
Effect of betaxolol (BTX, 5 mg/kg, i.p.) on established methamphetamine (MAP, 1 mg/kg, s.c.) induced CPP in mice. Data are presented as mean±standard error of the mean (each group N=8). *p<0.05, **p<0.01. Saline-MAP: saline was injected 24 h prior to the CPP testing session, BTX-MAP: BTX was injected 24 h prior to the CPP testing session. CPP: conditioned place preference.
Three days after the extinction of CPP, the priming injection of MAP (0.125 mg/kg, s.c.) caused a reinstatement of place preference. Figure 4 shows that the pretreatment of BTX (5 mg/kg, i.p.) significantly inhibited the reinstatement that was caused by the priming dose of MAP (p<0.001; Kruskal-Wallis test).
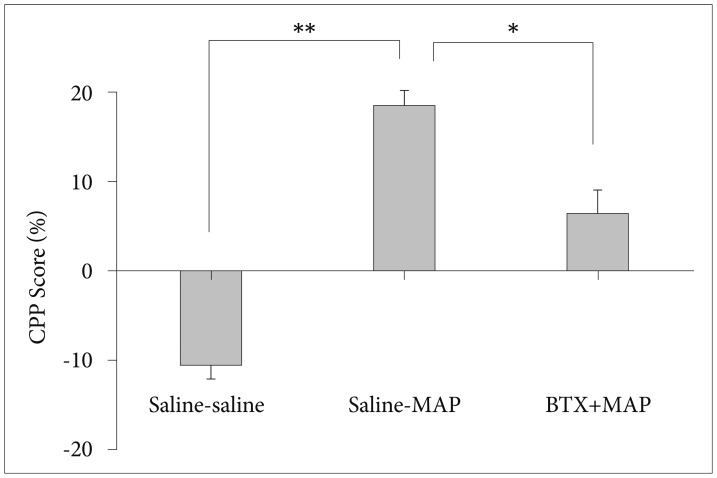
Effect of BTX (5 mg/kg, i.p.) on the reinstatement of CPP primed by methamphetamine (MAP, 0.125 mg/kg, i.p.) in mice. Data are presented as mean±standard error of the mean (each group N=8). *p<0.05, **p<0.001. Saline-MAP: saline injection 24 h prior to the priming injection of MAP, BTX-MAP: BTX injection 24 h prior to the priming injection of MAP. CPP: conditioned place preference.
DISCUSSION
In the present study, we examined the effect of the selective β1 antagonist BTX on MAP-induced CPP. To our best knowledge, the influence of BTX on the rewarding properties of MAP has not been investigated in a research study. In our results, we demonstrate the role of BTX in the development and expression of MAP-induced CPP and hyperactivity. The CPP paradigm was widely studied in rodents, which previously showed of a place preference for compartment paired with drugs such as MAP, morphine, or cocaine.1920 In our results, MAP induced a rewarding effect in the CPP paradigm while the locomotor activity was sensitized, which is also consistent with previous results.212223 We found that administering BTX just prior (30 min) to the administration of MAP suppressed the development of MAP-induced place preference; however, when administered 30 min prior to a MAP injection, BTX did not result in any changes of locomotor activity. Also, we failed to find a BTX-induced place-preference effect, indicating that BTX has no rewarding properties. This result could suggest that BTX may play a significant role in blocking the primary rewarding properties of MAP, which supports the evidence regarding the role of NE mechanisms in the addictive effects of psychostimulants. In contrast to acquisition, the expression of MAP-induced place preference occurred in the absence of the primary rewarding stimulus, for example MAP, and instead relied on the motivational properties of the conditioned stimuli, for example environmental cue. In addition, the administration of BTX 24 h prior to the CPP test day also attenuated the expression of any previously established MAP-induced place preference. Our result is consistent with previous studies that reported that the rewarding effects of psychostimulant drugs were decreased with the administration of a beta-adrenergic receptor antagonist; specifically, propranolol disrupted place preferences that were conditioned to morphine24 and cocaine.817 The present finding supports the evidence that the efficacy of BTX in psychostimulant abuse is potentially associated with its effect on a β1-adrenergic-receptor antagonism;25 however, the persistence of the BTX-induced attenuation of CPP for more than 24 h in our results remains unexplained.
Our result showed that a low dose of MAP (0.125 mg/kg), which produces no rewarding effects on its own, reinstated the extinguished place preference. However, we demonstrated that a MAP-induced reinstatement of CPP was significantly attenuated after pretreatment with BTX 24 h prior to the administration of MAP. Our findings are consistent with previous studies that showed that a morphine or cocaine injection reinstated an extinguished place preference;262728 therefore, our present finding further confirms that drug priming is an important factor that is implicated in relapse.
The illicit psychostimulant drugs–cocaine and the amphetamines, as well as their derivatives–represent a highly addictive class of compounds; among these, a surge in the popular use of MAP has occurred. The psychological effects of MAP use are associated with anxiety, depression, psychosis, and suicide.29 MAP has various neurotoxic effects including a persistent depletion of dopamine, an inactivation of the dopamine transporter, an inhibition of tyrosine hydroxylase, the dysfunction of the vesicle monoamine transporter, the degeneration of fine unmyelinated axons, and apoptosis.3031 MAP's neurotoxic effects on dopamine and serotonin neurons3233 are associated with the damage of brain regions such as the caudate, putamen, nucleus accumbens, substantia nigra, and hippocampus.34 In correspondence with an increased understanding of the mechanism of MAP use, a growing interest in the development of new pharmacological treatment options for treating MAP addiction and restoring neuronal function has emerged. We anticipate that our result may be elucidative with respect to these issues.
In conclusion, the present study suggests that the selective β1 adrenoceptor is involved in the reward-processing mechanisms of drug abuse; furthermore, the selective β1 adrenoceptor is also involved in the relapse to MAP craving. The selective β1-adrenoceptor inhibitor may be a novel pharmacological target for the management of MAP abuse and relapse prevention; however, research still needs to elucidate whether these post-retrieval mechanisms are permanent or transient.
Acknowledgments
This paper was supported by research funds of Chonbuk National University in 2011.