INTRODUCTION
Circadian rhythms are defined as bioquemical, physiological and behavioral variables, which show an approximately 24-hour cycle oscillation, reacting primarily to the light-dark environmental cycle.1 Preceding studies indicate they are found throughout wide-ranging species, including animals, plants and microorganisms.23 Chronobiology studies biological rhythms, of which circadian rhythms are an example.4
The central biological “clock” that controls circadian rhythms is located in the hypothalamus, consisting of a group of nearly 20,000 neurons named suprachiasmatic nucleus (SCN).2 Previous studies have already recognized genes that drive circadian rhythms in humans and numerous other species, which are frequently called circadian or “clock” genes.56 This group of genes are highly expressed in the SCN and their coordinated operation is responsible for the translation of the environmental circadian input into the organism.567
Circadian rhythms can influence sleep-wake cycles, hormone release, body temperature and other important bodily functions.3 They have been linked to various sleep disorders.8 Abnormal circadian rhythms have also been associated with obesity, diabetes, and psychiatric disorders such as Depression and Bipolar Disorder (BD).910
Some prior studies have revealed that, along with features connected to the sleep-wake cycle (chronotype, actigraphic measures), other variables related to circadian rhythms appear to be altered in BD. In fact, abnormalities of circadian rhythms such as body temperature, blood pressure, heart rate, hormone and neurotransmitters levels have been identified in patients with BD.1112 Circadian rhythms are important in determining human sleep patterns as well.13 Likewise, researchers have demonstrated that sleep disturbances are frequent between BD patients.14 Moreover, we have recently presented that poor sleep quality is a prevalent feature in BD patients even during the interepisodic phases of the disorder.15
Bipolar Disorder is a complex and multifactorial psychiatric disorder.16 Previous evidences from genetic research have confirmed that BD is highly heritable.17 Even though BD exhibits significant genetic vulnerability, the specific underlying genetic factors remains unclear.17 Growing amounts of evidences have implicated circadian genes in the heritability of BD. A number of genetic association studies have demonstrated associations between BD and various circadian genes, such as the Clock, Npas2, Artnl1, Per3 and Nr1d1 genes.18
The human PERIOD3 (Per3) gene is an important member of the Period family that was first isolated in mammals in 1998.19 It is located on human chromosome 1 (1p36.23) and is highly expressed in the suprachiasmatic nucleus.19 One study have recently showed that lithium, one of the first choice pharmacological agents for BD, alters the expression of the Per3 gene in laboratory cultures of fibroblasts.20 Genetic linkage studies of BD showed positive results for the chromosome 1 region where the Per3 gene is located.2122
Genetic association studies have shown some positive results of the Per3 gene and BD plus certain clinical subphenotypes, although there have been preceding publications that found no evidence of association between the Per3 gene and BD.2324252627282930 Thus, given the previous evidences of sleep/circadian disturbances as well as the genetic susceptibility for BD, we decided to investigate the possible link between the Per3 circadian gene and BD. The aim of the present study is to assess the association of some polymorphisms of the Per3 gene in a sample of BD patients.
METHODS
Study design/subjects/protocol
This report consists of a genetic association case (BD) vs. control study of the Per3 gene. Data collection took place from 2009 to 2013. There were 209 cases and 213 controls in our sample. We further subdivided our BD sample into “good sleepers” (PSQI ≤5) and “poor sleepers” (PSQI>5) according to the Pittsburgh Sleep Quality Index (PSQI) global score, and then we assessed genetic association of the Per3 gene with sleep quality in the BD group (cases).31
The PSQI is a subjective sleep quality instrument, widely used in clinical and research settings and validated to the Brazilian Portuguese.32 The instrument contains seven sub-components, which are relevant features of sleep quality, and generates a global score ranging from 0-21. Global PSQI scores >5 indicates poor sleep quality, whereas scores ≤5 suggests good sleep quality. In order to keep mood variations impact on sleep patterns in a minimum level, we picked BD cases fulfilling criteria for euthymia or remission.33 Euthymia was settled as a score lower than seven in both Hamilton and Young mood scales for the last month along with the lack of criteria for current depressive, mixed or hypo/manic episode by the DSM-IV-TR criteria.333435 The MINI-plus structured interview based on DSM-IV diagnostic criteria was used to set up for psychiatric diagnosis in the BD group.36 Psychiatric longitudinal follow-up evaluated by medical records were also taken into account to endorse accuracy of psychiatric diagnosis. Our sample of cases was composed by adult BD outpatients of Hospital das Clínicas-UFMG psychiatric service.
Controls were recruited from the general population. Inclusion criteria for the control group were an age ≥18 and the lack of criteria for psychiatric diagnosis by the MINI-plus, according to the DSM-IV.
Genetic analysis
We collected peripheral blood samples from cases and control groups. Genomic DNA was isolated with the saline method.37 DNA quantification was performed on spectrophotometer device, and subsequently, its concentration was adjusted to 50 ng/µL. Genotyping was achieved by Real-Time PCR (RT-PCR) and allelic discrimination was completed by Applied Biosystems (ABI) producer's protocol in the 7500 RTPCR System. The protocol used TaqMan® Genotyping Master Mix (Applied Biosystems, Foster City, CA, USA).
The Taqman® is a type of assay supplied by ABI (Applied Biosystems, Foster City, CA, USA) in the versions assay-ondemand and assay-by-design, consisting of two pairs of oligonucleotide primers: forward and reverse (oligonucleotide concentration in 900 µM), a marker dye VIC®-detects the presence of the allele 1 (markers with final concentration 200 µM), a second marker FAMTM dye-detects the presence of allele 2 (markers with final concentration of 200 µM). PCR reaction included an initial denaturation at 95℃ for 10 min followed by 50 cycles at 95℃ for 15 sec and 60℃ for 1 min. Every reaction contained 5.0 µL of mix, 0.1 µL of probe, 3.9 µL of deionized water and 60 ng of DNA. Ten percent of the samples were genotyped again at random, as a quality control approach, to ensure the exclusion of possible laboratory errors.
We picked single nucleotide polymorphisms (tagSNP's) that would provide a good cover of the whole Per3 gene using HapMap database (www.hapmap.org) and prior published studies. We further used 20 ng of DNA of each patient for five TaqMan SNP's genotyping assays (rs228697, rs228727, rs228729, rs707467, and rs10462020) according to the tutoring of Applied Biosystems.
Statistical analysis
Gender, genotype and allelic distributions were analyzed by chi-square or exact Fisher test using the software SPSS 20.0. Age was analysed by Mann Whitney non-parametric test. We took p-values of 0.05 or less as evidence of statistical significance. Adjusted residuals, which enables us to analyse the residuals in each cell of the cross tabulation, were performed using a cut-off point of +1.96 and -1.96.
RESULTS
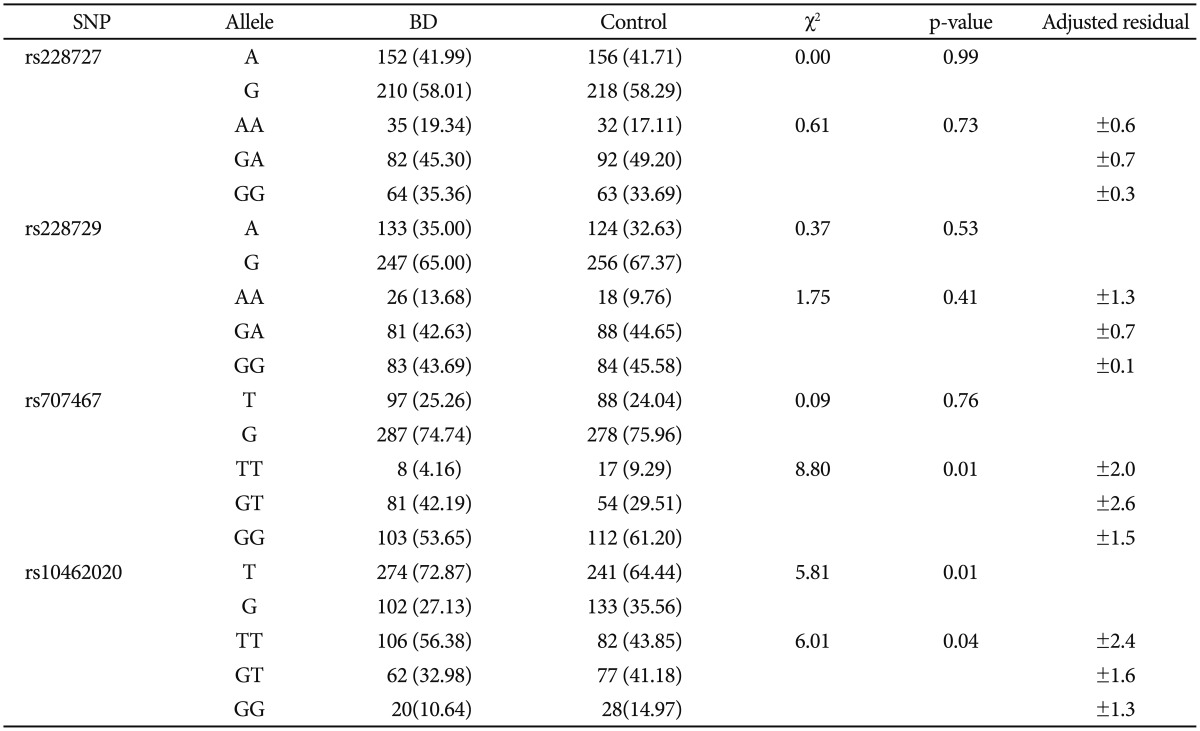
There were 209 cases and 213 controls in our sample. Groups were similar regarding age (Mann-Whitney U=22096.50; p-value=0.897) and gender (χ2=1.66; p-value=0.197). Median age were 43 for cases (min 18; max 88) and 42 for controls (min 18; max 86). 31.60% of cases were males, while 37.60% of controls were males. The SNP rs228697 failed to reach Hardy Weinberg equilibrium (HWE) and it was excluded of further analysis (χ2=17.07; p-value<0.01). All other SNP's were in HWE, except for rs10462020 which violated HWE as regards to the whole sample (χ2=7.20; p-value<0.01), although the control group was in HWE (χ2=1.92; p-value=0.16).
Table 1 shows the genotypic and allelic distributions of the Per3 gene SNP's assessed between cases and controls. The GT genotype of the SNP rs707467 significantly associated with BD (χ2=8.80; p-value=0.01; adjusted residual=±2.6). We also found significant association of the SNP rs10462020 allele T with BD (χ2=5.81; p-value=0.01) as well as the genotype TT with BD (χ2=6.01; p-value=0.04; adjusted residual=±2.4).
Table 2 shows the genotypic and allelic distributions of the Per3 gene SNP's assessed in the BD group subdivided into “good” and “poor” sleepers measured by the global PSQI score. No significant associations were found between sleep quality and the genotypic and allelic distributions of the Per3 gene SNP's in the BD sample.
DISCUSSION
The present study has limitations that should be well thought out when interpreting the results. Small sample size would be a major limitation and future studies should make efforts in achieving bigger samples of BD patients.
Several hypotheses can be thrown to try to understand the HWE deviations found in genetic association studies. Genotyping errors are one of the most common causes of deviation from HWE, usually resulting in increased number of heterozygotes from the expected.3839 In the other hand, when the number of heterozygotes are below the expected, one of the most likely possibilities to explain a HWE deviation is due to ethnic stratification and selection of the samples.383940 In our study, the HWE deviation of the SNP rs10462020 was due to a deficiency of heterozygotes than would be predicted by the HWE law.
The population stratification arises from differences between distinct ethnic groups.40 In addition, the Brazilian population presents considerable degree of miscegenation,414243 which could contribute to the HWE deviation found. Other hypotheses have also been thrown while trying to explain the HWE deviations eventually found in genetic studies.40 Some authors suggest that the deviation from HWE may be exactly an evidence of a genetic association.40 In other words, the deviation itself could accurately represent a possible role of the polymorphism in the predisposition to the disease assessed.444546
It has been suggested that the genetic vulnerability of BD would be of complex polygenic nature, with multiple susceptibility genes each conferring small effects.47 Therefore, the individual effect of each gene in determining the vulnerability to BD would be of minor magnitude and may not always reach statistical significance.47 However, in addition to the effect of each individual gene, other genetic factors may confer increased risk for BD, including structural genomic variations (CNV's), mitochondrial DNA variation and genetic and epigenetic interactions.47 In fact, some recent publication have demonstrated associations of epigenetic interactions between specific circadian genes, including the Per3 gene, and BD.4849 Thus, at least partially, the past mixed results of association between the Per3 gene and BD may be related to all these referred reasons as well as to the different compositions of the samples of each study, as mentioned in the last paragraph.
Still as regards to the mixed results of association between BD and the Per3 gene, previous studies showed that the regulation of the circadian system would be far more complex than initially proposed.5051 Consequently, in addition to the core clock genes directly involved in the central regulation of the circadian system (Clock, Artnl1, Cry1, Cry2, Per1, Per2, and Per3), new evidence suggest that hundreds of genes may play important role for the intricate process of the circadian regulation system.52 As a result, changes accumulated in a numerically larger set of circadian genes may possibly work in an integrated way towards an underlying vulnerability factor for BD.525354
In this sense, the use of new approaches in order to pool into the same analysis some of the genetic factors related to the vulnerability of BD can lead to advances in understanding the underlying susceptibility of the disorder.53 Convergent functional genomics and pathway analysis studies try to overcome this difficulty by integrating biological findings from various sources to genetic results arising from GWAS studies.525354 Therefore, instead of analyzing the association of a gene individually, which may frequently not reach the statistical significance threshold, these studies evaluate groups of genes involved in the same physiological pathway functionally related to the studied disease. Recently, some such studies that evaluated several genes involved in the regulation of the circadian system found positive association with BD.52535455 Thus, it is possible that these new methodologies will be able to better translate the polygenic nature of the genetic vulnerability to BD.
In addition to the limitations of association case-control studies involving a single candidate gene in identifying the underlying heritability of complex genetic disease such as BD, listed in the last paragraphs, some other limitations should be considered when evaluating the results of this study. The PSQI assesses sleep quality and its use is widespread both in the clinical setting and in scientific research.32 However, a major limitation of this study involves the low PSQI's ability to capture, in its full complexity, the various aspects of the socalled phenomenon of sleep-wake cycle. In fact, researchers claim sleep as one of the least understood topics in biological sciences.8 Furthermore, while evaluating sleep with the PSQI in the period related to the last 30 days, one cannot exclude the interference of a recall bias. Moreover, by capturing an average behavior, the instrument does not gather information regarding the variability of the sleep patterns day after day, which would be best captured by actigraphy or sleep diaries. Therefore, other phenomena related to the sleep function, which may be not fully captured by the PSQI, might be associated with the Per3 gene. However, despite sleep questionnaires are considered subjective sleep parameters, some evidence showed correlation of these with objective parameters measured by actigraphy and polysomnography in a sample from patients with BD.56
One last relevant issue that needs to be further addressed is the possible shared vulnerability of Sleep Disorders and BD. In fact, since sleep was recognized to be controlled genetically, researchers have showed that the mechanisms of sleep regulation and pathways, such as the circadian system and some neurotransmitter signaling, have conserved effects on sleep across a wide range of species up to humans.8 At the same time, genome-wide association studies (GWAS) have exposed a few loci rising vulnerability to primary Sleep Disorders as narcolepsy and restless leg syndrome.8 Thus, it is likely that, at least partially, Sleep Disorders and BD may join common liability factors.
Some positive aspects of this report should be highlighted. This is, to the best of our knowledge, the first report finding direct and significant association of the Per3 gene polymorphisms and BD in Brazilian and South American samples, as well, of BD patients. This should be of some relevance since Brazilian population has a large degree of miscegenation, as mentioned before, if compared with Caucasian populations. A previous study assessing a Brazilian sample of BD had found association of poor sleep quality and the Per3 gene, but not with BD itself.57 Another interesting aspect of our study is that we found evidence of association of BD with polymorphisms of a gene (Per3) considered a member of the core “clock” circadian system regulation. Additionally, the control group of our report did not differ significantly from cases (BD group) in clinical variables such as age and gender. This is important because researchers claim that one of the main causes of type 1 error in genetic association studies refers to issues regarding specific aspects of the control groups, such as poorly matched control groups.
In this study, we demonstrated evidences of genetic association between the Per3 gene and BD. Our results are consistent with recent findings showing association of the Per3 gene polymorphisms with improvement in depressive symptoms,58 with a family history of suicidal behavior,59 and with sleep disturbances in different samples of BD patients.48 Therefore, even taking into account the limitations of the current study, the results of association between the Per3 gene and BD in our sample may bring additional strength to the former evidences of association among the Per3 gene and BD. Furthermore, this finding is consistent with previous results of association of BD with other genes directly or indirectly related to the circadian regulation.60 Still, if considered together with all the scope of evidences implicating sleep and circadian disturbances with the fundamental pathophysiology of BD,59 as reinforced by recent results of convergent functional genomics and pathway analysis studies,52535455 our findings may enhance the core rationale connecting circadian regulation abnormalities to the neurobiology of BD.
In upcoming studies, assessing multiple control samples would minimize bias due to population stratification. Another interesting approach would be the use of prospective control samples, since generally these control groups are built retrospectively. Furthermore, given the multifactorial nature of BD, studies and techniques that provide the convergence of genetic association, gene expression and epigenetic data should be encouraged. Likewise, future efforts of researchers might focus in testing the shared impact of sets of genes related to the circadian regulation instead of testing one or a few genes individually, since it is reasonable to hypothesize that, according to the complex polygenic nature of BD, the underlying predisposition may be related to the disrupted function of several circadian genes which would, in turn, translate a disrupted circadian system regulation in BD.