INTRODUCTION
Evidence suggesting a strong association between immunological factors and the pathophysiology of major depressive disorder (MDD) has been consistently increasing today.1,2,3,4,5 A family history of autoimmune diseases has been also found to be significantly more frequent among patients with mood disorders, which suggests a possible relationship between the inheritance of immune disorders and mood disorders.6,7,8 However, the potential role of cytokines has not yet received sufficient attention despite their critical roles in the human immune system in relation to vulnerability to MDD.1,4,9,10,11 Among cytokines, monocyte chemoattractant protein-1 (MCP-1), a CC-chemokine, plays a very important role in the collection of monocytes and T-lymphocytes through regulating both the T helper-1 (TH-1) and TH-2 systems.12 MCP-1 is produced constitutively or after induction by oxidative stress, cytokines, or growth factors by a variety of cell types, including monocytes, smooth muscle cells, and endothelial cells.13 It is also expressed in highly regionalized neuronal areas such as the cerebral cortex, hippocampus, and hypothalamus, which are important in the pathophysiology of MDD.14 Additionally, MCP-1 expression is related to the development of various central nervous system (CNS) diseases, such as multiple sclerosis15 and Alzheimer's disease,16 which are commonly accompanied by depressive symptoms.7
In the context of evidence of a relationship between cytokines and the pathophysiology of MDD,17 MCP-1 is an interesting cytokine insofar as it is involved in alterations of the TH-1 and TH-2 systems. As MCP-1 may affect innate and adaptive immunity through monocyte regulation and the TH systems,18 it is plausible that MCP-1 may contribute to the development of MDD, which would support the TH-1/TH-2 hypothesis.19
Evidence of the Potential Role of Monocyte Chemoattractant Protein-1 in the Development of MDD
Neuroanatomical basis
A recent animal study found that MCP-1 may act as a modulator of neuronal activity and neuroendocrine functions given that the expression of MCP-1 mRNA and protein is regionalized in the CNS in neuroanatomical sites implicated in the development of MDD14,20 (e.g., the cerebral cortex, hippocampus, paraventricular and supraoptic hypothalamic nuclei, lateral hypothalamus, substantia nigra, and cerebellum).
Co-localization and interaction with crucial biological molecules associated with MDD
Other evidence comes from MCP-1's co-localization with neurotransmitters and neuropeptides in well-defined brain nuclei, the so-called cholinergic, dopaminergic, vasopressinergic, and melaninconcentrating hormone (MCH)-expressing neurons,14 which are primarily involved in the symptomatology and therapeutic mechanism of MDD.21 For example, noradrenaline induces production of MCP-1 in astrocytes and thereby mediates neuroprotective actions.22 Additionally, according to a recent animal study investigating a relationship between MCP-1 and stress, exposure to stress produces a moderate increase in MCP-1 through the inhibitory activity of glucocorticoids during the stress reaction. Likewise, the corticosterone treatment of astrocytes significantly reduces production of MCP-1. When the production of glucocorticoids by stressed rats was blocked by the selective inhibitor metyrapone, a large increase in the concentration of MCP-1 was observed in the cortex. Desipramine, one of the noradrenergic antidepressants, was found to increase MCP-1 in the cortex. These data suggest that MCP-1 may play a critical role in altering noradrenaline or glucocorticoids, which may be implicated in the development and treatment of MDD.23 Additionally, the presence of MCP-1 in magnocellular vasopressinergic neurons raises the possibility that MCP-1 may contribute to the control of eating and drinking behaviors, which strengthens the plausibility of the notion that it could exert neuroendocrine functions.14 In a recent study investigating a MCP-1 level in cerebrospinal fluid (CSF),24 137 psychiatric patients with a history of a suicide attempt and 43 healthy controls was investigated. In the study, a subgroup of patients (n=42) was followed up with blood samples 12 years after the initial CSF sampling, when they did not show suicidal behavior. The follow-up chemokine levels were compared to those of psychiatric patients (n=17) who had never attempted suicide. According to the results, the MCP-1 levels were significantly lower in the suicide attempters than in healthy controls. In addition, patients with depression had the lowest CSF levels of MCP-1, although no correlations were found between the level of MCP-1 and severity of depression which was measured by the Montgomery-Asberg Depression Rating Scale. The findings suggest that the exact role of MCP-1 in the context of pathophysiologies associated with suicidal behavior and depression should be further investigated.24
Interaction with cytokines
Moreover, MCP-1 has an important role in the modulation of other cytokines. For example, it stimulates the expression of interleukin (IL)-4 in the TH-2 system,18,25 which is consistently reported to be altered in patients with MDD.26 It should also be noted that IL-4 is involved in the antidepressant treatment response27 as well as in the regulation of IL-6 and IL-8 (both ILs are altered in MDD).18,27,28
Genetic studies
Genetic susceptibility to MDD may be explained by the presence of specific polymorphisms in chemokine genes, leading to abnormal protein expression or functioning.29 It has also been noted that the MCP-1 gene is located on the chromosome 17q11.2-q12 (1.5 kb in length and containing three exons and two introns).30 Intriguingly, this chromosomal region is adjacent to the encoding area for the serotonin transporter gene (5-HTTLPR),31 which is one of well-known candidate genes for MDD that has a promising pharmacogenetic profile in relation to antidepressant efficacy. Two polymorphisms in the promoter region of the gene (A-2518G and A-2076T) have been identified.32 The A-2518G polymorphism affects the transcriptional activity of the distal regulatory region and monocyte MCP-1 production.32,33,34 Recent data have indicated that MCP-1 gene -2518 polymorphism (allele A) was associated with an increased risk for MDD [odds ratio (OR)=1.83, 95% confidence interval (CI)=1.18-2.83], which suggests that the MCP-1 gene -2518 polymorphism may confer susceptibility to MDD.35 A gene-dose effect was also found when the genotypes AA and AG were individually compared with the genotype GG (p for trend=0.008): 1) AA vs. GG, p=0.02, OR=3.06, 95%CI=1.21-7.83; 2) AG vs. GG, p=0.21, OR=1.56, 95%CI=1.56-3.00.35 In a subsequent preliminary study with small MDD (n=34) and bipolar disorder (BD, n=61) samples and a healthy control group,33 no significant association between the A-2518G polymorphism of the MCP-1 gene and any type of diagnosis was found. However, when the different diagnostic subtypes (MDD vs. BD) were considered, numerical superiority of the AA genotype and of the A allele of the A-2518G polymorphism of the MCP-1 gene was observed in subjects with BD.33 Moreover, more suicide attempts in subjects with the AA genotype of the A-2518G polymorphism of the MCP-1 gene were also observed in this study.33 Additionally, a significant association between BD and suicide attempts has been identified in patients with this disorder, who are also more likely to have the AA genotype of the A-2518G polymorphism of the MCP-1 gene. Furthermore, the MCP-1 gene -2518 polymorphism was also consistently associated with the development of BD and schizophrenia,33,34,36 although another study failed to replicate the association between the MCP-1 gene polymorphism and bipolar disorder.37
Neuroprotection, including the oxidative stress pathway
Oxidative stress occurs when the redox balance is breached and pro-oxidative processes overwhelm the antioxidant defense system in the brain. Factors favoring oxidative processes may also have roots in genetic endowment, immune activation, psychosocial stress, or energy failure (e.g., stroke).38,39,40,41 Nitric oxide (NO) is a ubiquitous neurotransmitter involved in a various brain functions and neuropathologies (e.g., dopamine, norepinephrine, and glutamate release and neuronal cell survival) through modulation of oxidative stress-related sequential cascades in the brain. Inflammation or neuronal excitation leading to increased intracellular Ca2+ may enhance production of NO. It has been suggested that optimal modulation of NO is also crucial in the development of MDD.42 Along with immune activation, neuronal toxicity may also contribute to oxidative stress. Proinflammatory cytokines have been reported to enhance indoleamine 2,3-dioxegenase (IDO) activity43,44 under stress, promoting the kynurenine pathway instead of the 5-hydroxytryptamine (5-HT) pathway of tryptophan.45 As a result, 5-HT synthesis is reduced, and quinolinic acid, known to be neurotoxic,46 is increased. MCP-1 has also been associated with the activation of microglia,47 probably initiating the release of NO. According to a recent in vitro study,48 noradrenalin induces expression of MCP-1 in astrocytes, which is neuroprotective against excitotoxicity in neuronal cells. In primary neurons, MCP-1 reduced NMDA-dependent glutamate release, glutamate-dependent Ca2+ entry, adenosine triphosphate (ATP) loss, and LDH release attributable to NMDA or glutamate. MCP-1 also reduced the toxic consequences of oxygen-glucose deprivation in neurons, as did conditioned media from astrocytes, which could be prevented by a blocking antibody for MCP-1. These findings indicate that the neuroprotective effects of noradrenaline may be partly mediated by the induction and release of astrocyte MCP-1.48
Excitatory neurotransmitters may cause neuronal death via multiple synergistic injury mechanisms in the CNS.49 Of note, one such mechanism is also initiated by microglia and the microglial chemotaxis that can be signaled by MCP-1, leading to excitotoxic neurodegeneration.49 The hippocampus, which is critical for mood and emotional regulation, is particularly vulnerable to the excitotoxic brain injury associated with MCP-1.50 The aforementioned findings underscore the importance of MCP-1 in modulating certain processes within the cascade of excitotoxic insults consistently suggested as a possible etiological factor in the development of MDD.51 Furthermore, MCP-1 is associated with the regulation of glial cells, resulting in alterations of neuroprotective substances and nerve growth factors (NGFs) in the CNS, which suggests that MCP-1 plays a role in regulating neuronal cell function through an indirect mechanism involving the modulation of glial cells.52
Involvement in comorbid disorders
Associations between MCP-1 and various CNS diseases that are frequently accompanied by comorbid depressive symptoms, such as multiple sclerosis,15 stroke,53 and Alzheimer's disease,16 have also been suggested. Interestingly, plasma MCP-1 levels were found to be significantly in patients with chronic renal failure associated with coronary heart disease, who are especially vulnerable to the development of MDD, compared with controls.54,55 A limited number of studies have also suggested that patients with MDD without conventional risk factors for coronary heart disease may have higher levels of MCP-1 than do healthy controls.55
Response to antidepressants
Intriguingly, a recent study that investigated the role of immune dysfunction in MDD (n=40)56 found that patients with MDD showed clear evidence of immune alteration; they had higher levels of IL-6 and IL-10 but lower levels of IL-4 in the context of a hypothalamic-pituitary-adrenal axis (HPA) disturbance characterized by higher cortisol levels. In this regard, evidence indicating that the alteration of MCP-1 levels as a biomarker for the development of MDD remains sparse, suggesting that research investigating the role of MCP-1 as a crucial molecule for MDD is still in its infancy. Nonetheless, a recent human study56 found that patients with MDD refractory to antidepressant treatment showed significantly lower levels of MCP-1 levels than did responsive patients at baseline, although MDD patients did not differ from healthy controls in terms of MCP-1 levels. This finding is the first to reflect an association between MCP-1 and the outcome of antidepressant treatment.56 These data suggest that the lack of a therapeutic benefit from antidepressants may, in part, be associated with alterations in MCP-1 levels. In support of these findings, reported by Carvalho and colleagues, studies examining antidepressants and noradrenergic activity have also demonstrated that antidepressants may have the potential to down-regulate chemokines such as MCP-1 in MDD, suggesting that MCP-1 may be a candidate for a new pharmacological target.57
The Table 1 summarizes the potential role of MCP-1 in the development and treatment of MDD.
Areas for future research
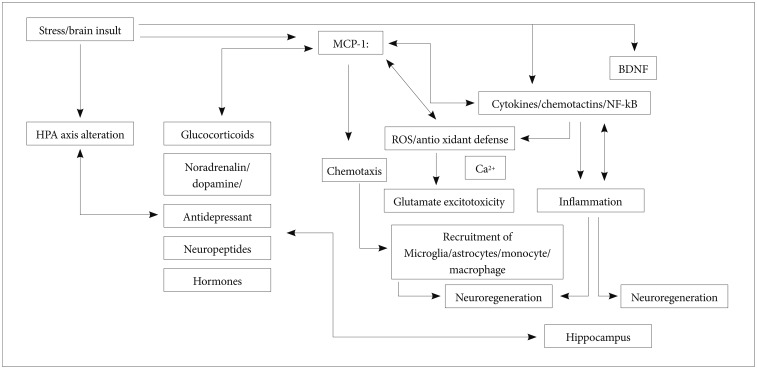
The possible role of MCP-1 in the development of MDD deserves further exploration, even though currently available findings do not definitely indicate that MCP-1 is directly implicated in the development of MDD. However, it is likely that MCP-1 modulates neuronal activity and neuroendocrine functions such as MCP-1 mRNA and protein that have been highly selectively found in MDD-related CNS regions such as the cerebral cortex and hippocampus. Moreover MCP-1 is colocalized with cholinergic, dopaminergic, vasopressinergic, and melaninconcentrating hormone (MCH)-expressing neurons, which are critical in the development and treatment of MDD. MCP-1 may also regulate the pivotal step in the communication among crucial cytokines such as IL-4, IL-6, and IL-8, which are involved in the pathogenesis of MDD. Additionally, MCP-1 may be a possible modulating factor intervening in the process of excitotoxicity and neurodegeneration. Finally, new evidence that MCP-1 is implicated in the treatment response to antidepressants is also emerging, although this issue should be extensively and further investigated in future well-designed and adequately powered studies. Hence, we can plausibly speculate that MCP-1 plays a potential role in the etiopathogenesis of MDD. The Figure 1 illustrates a schematic involvement in MDD.
CONCLUSION
MCP-1 is a chemokine that may play a major role in MDD given its association with key cytokines/neurotransmitters/neuropeptides. Indeed, it has been found to influence the regulation of neuronal activities involved in the susceptibility to MDD. It is plausible that investigation of the role of MCP-1 in the development of MDD will provide new insights into the pathogenesis of this complicated and heterogeneous CNS disease. Moreover, co-localization of MCP-1 mRNA and protein with monoaminergic and neuropeptide neurons constitutes critical information relevant to the development of newer antidepressants. The proposed pathophysiologies of MDD are multifactorial, and aberrations in neurotrophic factors, alterations in neurotransmitter-receptor signal pathways, disturbances in the HPA axis, inflammation, immune dysfunctions, imbalance between oxidative stress and antioxidant defenses, and neuroprotective and mitochondrial dysfunctions have been consistently hypothesized as major pathophysiological processes in the development of MDD. Therefore, data implicating MCP-1 in multiple pathways involved in the development of MDD may also contribute to the identification of possible new pharmacological targets for the treatment of the heterogeneous psychiatric disorder known as MDD.