INTRODUCTION
Mood disorders are among the most prevalent forms of mental illness. Depression is a widespread and heterogeneous disorder with severe health and socioeconomic consequences.1 The monoamine hypothesis of depression, which includes dysfunction in the serotonergic, noradrenergic and possibly dopaminergic systems, is widely accepted as the pathophysiology of depression. The introduction of selective serotonin reuptake inhibitors (SSRIs) and serotonin-norepinephrine reuptake inhibitors (SNRIs) gave clinicians a safer medication choice than tricyclic antidepressants (TCAs).2 However, the efficacy of SSRIs and SNRIs is not better than that of TCAs, suggesting the need for rational drug discovery of antidepressants. Tianeptine is a newer antidepressant with unique neurochemical properties that are different from other antidepressants.3 In contrast to TCAs and SSRIs, tianeptine is suggested to decrease serotoninergic activity and the amount of serotonin in serotonergic synapses by increasing serotonin reuptake; it is hence called a serotonin reuptake enhancer.4,5,6 Clinically, tianeptine has an efficacy equivalent to the classic antidepressants.7
Tianeptine decreases the axonal atrophy of neurons that is precipitated by stress and prevents the loss of spatial memory induced by chronic stress and the cognitive deterioration that results from drinking alcohol.8,9 Shortening of dendritic length in the pyramidal neurons of the CA3 region of rats exposed to stress was observed only in a group treated with tianeptine, and not in a group treated with fluoxetine.10
Depression is considered to be caused by structural changes in molecules such as monoamines in the brain, and antidepressants function by counteracting these molecular changes. However, several observations suggest an alternative hypothesis to this view of depression.11 According to the network hypothesis, depression results from an abnormality of information processing within neural networks, and antidepressant drugs improve the plasticity of neuronal connectivity.12 Experimental studies of stress and antidepressant treatment implicated neurogenesis in the etiology of depression. Antidepressant drugs increase the regeneration and turnover of neural cells to replace apoptotic cells with new viable cells. Antidepressants also induce neural plasticity by contributing to the maturation of new neural cells.11,13
In general, the improvement of depressive symptoms by long-term antidepressant administration is attributed to the restoration of neural plasticity.14 One of the indexes of the restoration of neural plasticity is neural cell adhesion molecule (NCAM). NCAM is a cell-surface protein that contains five Ig-like and two fibronectin type III repeats followed by a variably spliced cytoplasmic domain that produces two major transmembrane isoforms (180 and 140 kDa) and a glycophosphatidyl inositol-linked isoform (120 kDa). The cytoplasmic domains of the NCAM isoforms link to the actin cytoskeleton by directly binding to spectrin and also interact with integrin to activate signaling pathways.15 The homophilic interaction of NCAM promotes the binding of Fyn (a nonreceptor tyrosine kinase) and Fak (a focal adhesion kinase) and their phosphorylation, which activates the Ras-mitogen-activated protein kinase (MAPK) pathway. The cyclic-AMP response element binding protein (CREB) is subsequently phosphorylated, resulting in neurite outgrowth. Consequently, NCAM promotes survival and differentiation of neurons, axon growth, and synapse formation.16,17
Among the three NCAM isoforms, the expression level of NCAM140 is sensitively regulated by chronic stress. Downregulation of NCAM140 is implicated in the structural alterations of the hippocampus induced by chronic stress exposure.18 In addition, NCAM140 has protective activity against oxidative stress.19
NCAM140 is known to be important in the formation and maintenance of the neural network. However, little is known about the role of NCAM140 in antidepressant action. Previous studies reported that fluoxetine affects NCAM140 expression.20,21 The aim of this study was to identify the effect of tianeptine treatment on expression of NCAM140 and downstream signaling molecules. To explore this, we measured the expression of NCAM140 and CREB/phosphor-CREB (pCREB) in SH-SY5Y cells after tianeptine treatment. We identified significant upregulation of NCAM expression and phosphorylation of CREB in tianeptine-treated SH-SY5Y cells. These results suggested that the antidepressant effect of tianeptine might result from the regulation of NCAM expression and CREB phosphorylation.
METHODS
Cell culture
SH-SY5Y, a human neuroblastoma cell line, was cultured in Dulbecco's modified Eagle's medium (DMEM) (Gibco BRL) containing 10% FBS (Gibco BRL). DMEM culture solution was changed every 48 hours until cell maturation. For experimental treatment, tianeptine (Sigma-Aldrich) was prepared at 10 or 20 µM before addition to cell cultures.
Protein separation and measurement
Protein separation was performed using the iNtRON Biotechnology kit. First, 10 or 20 µM tianeptine was added to SH-SY5Y cells. After 6, 24, or 72 hours of culture, cells were washed with PBS and harvested by 2000-3000 rpm centrifugation. To 5×106 SH-SY5Y cells, 400 µL Pro-Prep was added with mixing and cells were incubated on ice for 20 minutes. After 13,000 rpm centrifugation for 5 minutes at 4℃ , proteins in the upper layer were collected and stored at -20℃ for Western blotting and quantitative analysis. Protein assay kit reagents (Bio-Rad) were diluted 1:5 with distilled water, and 1, 0.5, 0.1, 0.01, or 0 mg/mL BSA standards were set up. After placing 10 µL of BSA standards in 96-well plates, 200 µL working solution was added. Prepared BSA standards and protein samples to be measured were placed at room temperature for 5 minutes. Absorbance was measured with an ELISA reader at 595 nm, and protein concentration was measured using the standard curve.
Western blot analysis
To observe NCAM140, CREB and pCREB, Western blots were performed with protein extracted from cells using polyclonal rabbit anti-NCAM (1 : 10,000) (Chemicon), anti-CREB or anti-pCREB (1 : 1,000) (Cell Signaling Technology). Protein samples were boiled for 5 minutes in Laemmli sample solution (62.5 mM Tris-Cl pH 6.8, 2% SDS, 10% glycol, 0.5% β-mercaptoethanol, 10 µg/ml bromophenol blue), and 80 V electrophoresis was carried out using 8% or 12% polyacrylamide-gels, and proteins in 20% methanol were electrotransferred at 30 V at 4℃ overnight to nitrocellulose membranes. Membranes were incubated in TBST buffer (20 mM Tris-HCL pH 7.6, 500 mM NaCl, 0.1% Tween 20) containing 5% nonfat dry milk and stirred at room temperature for 1 hour. Respective antibodies were added with stirring at 4℃ in TBST buffer overnight. Membranes were washed with TBST buffer 3 times for 10 minutes each, then washed with TBST buffer containing 1:2000 HRP-conjugated anti-rabbit IgG. ECL+reagents (Amersham Bioscience) were added at 1:40 and membranes were exposed to film. Experiments were repeated 6 times for data validation.
RESULTS
In a previous study, administration of 10 µM tianeptine had a neuroprotective effect on damaged neuronal cells.22 Hence, we used 10 µM tianeptine as an appropriate effective dose and 20 µM as a higher effective dose. To evaluate the viability of SH-SY5Y cells after tianeptine treatment, we incubated SH-SY5Y cells in media with 0, 10 or 20 µM tianeptine and maintained cells for up to 3 days. The proliferation and morphology of SH-SY5Y cells was not affected by tianeptine (data not shown). We therefore treated cells with 0, 10 or 20 µM tianeptine for 72 hours.
Dosage effect of tianeptine on NCAM140 expression
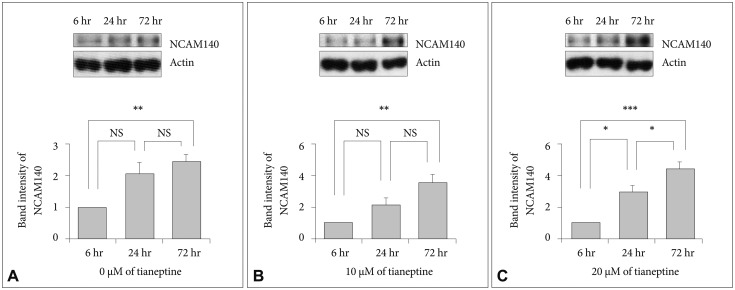
We explored the possibility that the increasing doses of tianeptine affected NCAM140 expression in SH-SY5Y cells. Tianeptine was administered to SH-SY5Y cells for 6, 24, or 72 hours at 0, 10, or 20 µM. The effects of tianeptine treatment on NCAM expression in SH-SY5Y cells were measured using immunoblotting (Figure 1). In 0 µM tianeptine-treated cells, NCAM140 expression increased by more than 105% at 24 hours and significantly increased by almost 143% at 72 hours compared to cells treated with tianeptine for 6 hours (Figure 1A). In 10 µM tianeptine-treated cells, NCAM140 expression increased by more than 112% at 24 hours and significantly increased by 254% at 72 hours. NCAM140 expression significantly increased by more than 190% at 24 hours and by 439% at 72 hours in 20 µM tianeptine-treated cells compared to at 6 hours. Moreover, NCAM140 expression after 20 µM tianeptine treatment increased earlier and to higher levels than after 10 µM tianeptine treatment. These results suggested that the tianeptine effect on expression of NCAM140 might be dependent on dose and length of administration.
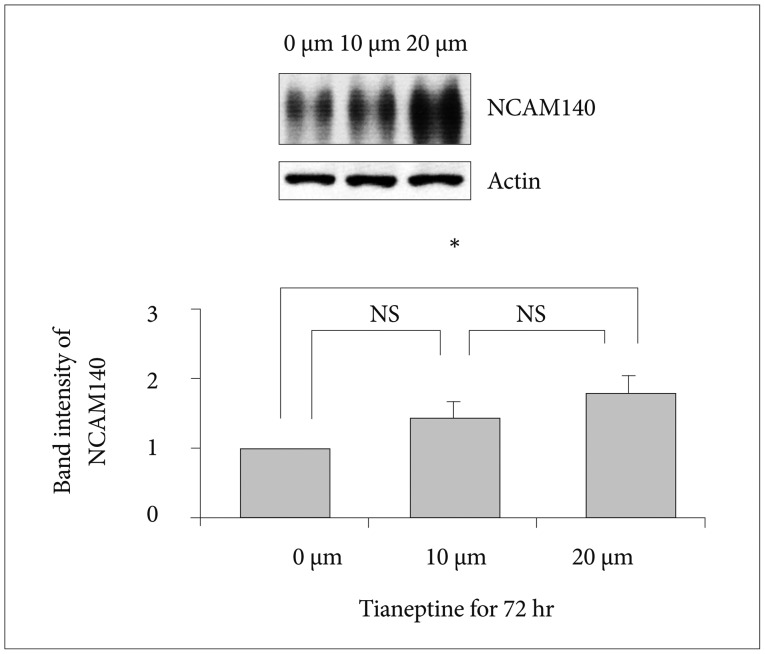
To explore the expression levels of NCAM140 at 72 hours, we administered 0, 10, or 20 µM tianeptine to SH-SY5Y cells and performed immunoblotting (Figure 2). We found that 20 µM tianeptine treatment resulted significantly increased NCAM140 expression compared to 0 µM treatment. In cells treated with 10 µM tianeptine, the expression level of NCAM140 was not significantly different than cells treated with 0 µM tianeptine but expression showed a tendency to increase. These results suggested that administration of tianeptine induced expression of NCAM140.
Effect of tianeptine on CREB expression and phosphorylation
Previously, fluoxetine treatment was found to activate NCAM140 expression and CREB phosphorylation in rat C6 glioma cells.21 We explored whether tianeptine treatment affected expression and phosphorylation of CREB. SH-SY5Y cells were treated with 20 µM tianeptine for 6, 24, or 72 hours and CREB and pCREB expression was measured by immunoblot (Figure 3). The CREB expression level at 24 or 72 hours was similar to the level at 6 hours. However, CREB phosphorylation gradually increased with time. These results indicated that tianeptine treatment seemed to activate CREB phosphorylation without affecting the total CREB level.
DISCUSSION
In this study, we found that cells treated with tianeptine for 72 hours showed a significant increase in NCAM140 expression compared to cells treated for 6 hours. In particular, 20 µM tianeptine for 24 hours induced a significant increase in NCAM140 expression in contrast with 0 or 10 µM tianeptine treatment (Figure 1). NCAM140 expression seemed to have a dose-dependent effect because 20 µM tianeptine for 72 hours significantly increased NCAM140 expression compared to 0 µM tianeptine and 10 µM tianeptine tended to modestly increase NCAM140 expression compared to 0 µM tianeptine (Figure 2).
Chronic treatment with imipramine, a tricyclic antidepressant, is reported to induce an increase in PSA-NCAM in the rat prelimbic cortex.23 Fluoxetine treatment of rat C6 glioma cells increases NCAM140 expression in a time-dependent manner.21 In contrast, a previous study reported that venlafaxine, an SNRI, attenuated the NCAM increase in the hilus area of the mouse hippocampus.24 Thus, the regulation of NCAM140 expression might be important in depression treatment.
Previous studies on the antidepressant effect and action mechanism of TCAs and SSRIs demonstrated that longer treatment of antidepressants is more important than any other condition of treatment.25 Some studies compared antidepressant and non-antidepressant treatment in rodents exposed to repeated chronic stress.26,27 Other studies measured the level of molecular markers representative of synaptic plasticity with diverse antidepressants treatment.28,29 The findings of these studies demonstrated that chronic stress exposure, which can precipitate or exacerbate depression, disrupts neuroplasticity. Antidepressant treatment has an opposing effect and can enhance neuroplasticity including structural plasticity changes in dendritic morphology and adult neurogenesis, and physiological synaptic plasticity. Chronic administration of a TCA blocked the decrease in spine density in the CA3 and CA1 cell layers of dentate gyrus in the hippocampus.30 Interestingly, chronic stress decreases the number and length of apical dendrites of CA3 pyramidal cells in the hippocampus.31 This reduction in dendritic length and complexity is blocked or reversed by chronic administration of tianeptine, but not by administration of a fluoxetine.10 Our data suggested that treatment of depressed patients with tianeptine might modulate the neural network by upregulating NCAM140 expression and resulting in improved mood.
Previous data demonstrates that NCAM induces recruitment of the tyrosine kinase Fyn to the NCAM intracellular domain, which in turn activates Fak, ERK, and the transcription factor CREB, which stimulates neurite outgrowth.32 CREB transcriptional activity depends on its phosphorylation state, with phosphorylation resulting in the activation of cAMP response element-mediated gene transcription and neuronal remodeling such as neurite outgrowth.33 In this study, CREB phosphorylation increased time-dependently with 20 µM tianeptine treatment (Figure 3). Future studies are needed to determine whether tianeptine treatment has an impact on modulating neuronal plasticity and neurite outgrowth by phosphorylating and activating CREB via NCAM140-induced activation of the Ras-MAPK pathway.
Increasing evidence suggests that the glutamatergic system, which is known to play a major role in neuronal plasticity and cellular resilience, may be involved in the pathophysiology and treatment of depression.34 In addition, the network hypothesis states that depression is defined as a disorder with impairment in information processing with structural changes in the brain.12 In this hypothesis, increased glutamate induced by stress might result in cellular excitotoxicity in the brain.35 Tianeptine treatment normalizes impaired glutamatergic neurotransmission in the hippocampus and amygdala induced by stress. Furthermore, tianeptine modulates excitatory synaptic currents of the hippocampal CA3 synapses to regulate the phosphorylation state of glutamate receptors in chronically stressed rodents36 and increases basal transmission of glutamatergic excitatory synapse.37 Finally, facilitation of AMPA receptor-mediated transmission leads to activation of the NCAM promoter, indicating that that NCAM synthesis is regulated in part by synaptic activity.38 Thus, tianeptine might induce NCAM140 expression by modulating glutamatergic transmission.
Our findings must be reviewed in the context of the study's limitations. We were unable to exclude that the antidepressant might regulate other isoforms of NCAM in addition to NCAM140. Experiments were performed only a single time for pCREB immunoblot data, so we were able to determine statistical significance. Additional studies are needed to establish these findings. We were unable to demonstrate the impact of the serotonergic modulating function of tianeptine on expression of NCAM140 and pCREB. We will investigate this mechanism in the future.
According to our findings, tianeptine has a distinct mechanism that is unlike other antidepressants. Therefore, studying the effects of tianeptine in patients with depression who are not adequately treated by SSRIs would be clinically informative. In conclusion, our study suggests that tianeptine has antidepressant effects through increased expression of NCAM140 and phosphorylation of CREB.