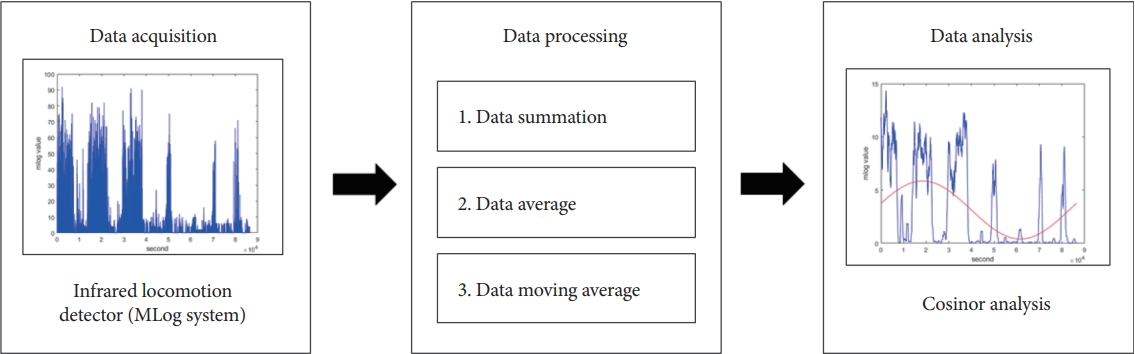
INTRODUCTION
Locomotor activity is one of the major assessments that reflects individual animal gross behavior in animal research [1,2]. Numerous research using locomotor activities have been performed to explore the changes of gross motor activities by experimental drugs [3]. In addition, using the strong association between locomotor activity and sleep-wake state, there were studies to investigate sleep-wake states [4]. Furthermore, locomotor activities can be analyzed in a perspective of circadian rest-activity rhythm [5,6]. Even though locomotor activities show the overall results of individual behavior that respond to the external situation at that moment, locomotor activities are bound to be under the gross influence of day and night. For this reason, locomotor activities can show circadian rest-activity rhythm that is one of the indices of circadian rhythms [7].
In the area of research of circadian rhythm, several studies have used the circadian indicies of rest-activity rhythm that involved locomotor activities. Among various methods for analysis of circadian rest-activity rhythm, cosinor analysis is a common analytic methodology in circadian rhythm research areas using locomotor activity [8]. Many studies have widely used cosinor analysis to analyze circadian rest-activity. However, specific analyzing methods such as sampling rates and data processing in raw data have been performed in various way [9]. In previous studies, sampling rates have been used at various time intervals from one minute to one hour. One minute can be used as a basic sampling rate [10,11]. Sampling rates in multiples of three, such as 3 minutes [12,13] or 6 minutes [14] can be widely used, because it can be easily interpreted visually considering that a day is 24 hours. On the other hand, there are common cases where it is summarized in multiples of five, such as 5 minutes [15,16], 10 minutes [17,18], or 15 minutes [5]. Meanwhile, raw data may be processed in various methods, such as data summation [3,19], data average [20-22], or data moving average [23].
Although cosinor analysis is performed in various ways in each study, it is necessary to provide sufficient rationality for these specific methods [9]. However, due to several reasons, there are many cases that are used without reasonable evidence. Although high-resolution information has recently been obtained due to the development of technologies for measuring locomotor activity [24], there are some opinions that such high-resolution information may not necessarily be good [24]. It depends on the research purpose, but in some studies, high throughout may act as noise. Therefore, an appropriate sampling rate is required according to the research purpose. As such, an optimal analysis method needs to be presented, and it is necessary to explore the relationship between these methods in order to compare existing studies. However, the difference among these data processing methods is unknown yet, and there was a lack of evidence for optimal analysis of circadian rest-activity rhythm. Therefore, we aimed to investigate the optimal strategy about the data processing method and the appropriate sampling rate for cosinor analysis in mice through a mathematical simulation according to the time block of data processing.
METHODS
Study subjects and procedure
Twenty male ICR mice (age: 4-5 weeks, weight: 21-27 g; Koatech, Pyeongtaek, Republic of Korea) were used in this study. The subjects were allowed to drink water and eat food freely. The temperature and humidity of the laboratory environment were maintained at 20°C±1°C and 40%-60%, respectively. The mice were acclimatized to the laboratory environment for a week. After the acclimatization period, the locomotor activities of the animals in homecages were collected every second by infrared red motion detectors for 24 hours. The illumination was controlled at the 12 h light-12 h dark cycle by an illumination control system (iW Blast Powercore/Colorplay 3/Data Enabler Pro; Philips, Burlington, MA, USA). The lights were set to turn off at 5 P.M. and turn on at 5 A.M. The Animal Experiment Ethics Committee of Pusan National University Hospital approved this study (IRB number: PNUH-2017-118).
Assessment of homecage locomotor activities
The locomotor activities in homecages were measured by the MLog system (Biobserv Inc., Bonn, Germany). The MLog system installed in homecage detects the free locomotor activity using infrared motion sensors. The MLog system data were collected every second. If there was no activity, it was recorded as zero, and more activities were indicated by higher data values.
Analysis and mathematics simulation
Circadian rest-activity rhythms were analyzed by cosinor analysis. Cosinor analysis is a type of analysis that finds an optimal cosine curve to show the least square of the difference between the fitted cosine curve and the original raw data [25]. Cosinor analysis estimates a circadian rhythm for 24 hours through a zero-amplitude test. The null hypothesis of zeroamplitude is that the amplitude is zero. Through cosinor analysis, circadian indices such as the amplitude, MESOR, and acrophase were calculated.
In order to find the most optimal time block for cosinor analysis, mathematical simulations were performed as explained in Figure 1. According to the increase in processing time per second in data summation, data average, and data moving average, the changes of circadian indices were simulated. When simulating the changes of circadian indices, the F and p values were calculated by zero-amplitude test to verify the accuracy of the values estimated by cosinor analysis. Cosine analysis, zero-amplitude test, and mathematical simulation were performed using MATLAB R2020b (MathWorks, Natick, MA, USA). Two values estimated by data average and data moving average were compared by intraclass correlation (ICC) and Pearson’s correlation by IBM SPSS version 22.0 (IBM Corp., Armonk, NY, USA), with a statistical significance level of less than 0.05.
RESULTS
Characteristics of estimated circadian rest-activity rhythms according to the methods of data processing
The estimated values of circadian rest-activity rhythm by data summation, data average, and data moving average are shown in Table 1. When using data summation, the estimated values gradually increased according to size of the time block. Meanwhile, the estimated values by both data average and data moving average were similar independent of time block. Based on the values estimated by data moving average, the amplitude was 3.631 (±1.137), MESOR 4.918 (±1.370), and acrophase -1.561 (±0.402), respectively.
Estimation of circadian rest-activity rhythm using data summation
When using data summation, the estimates of amplitude and MESOR gradually increased (Figure 2A). The acrophase showed a small variance within about 6,000 seconds, but the values were gradually increased according to the data processing time after 6,000 seconds. The F and p values in zero-amplitude test were statistically significant within about 800 seconds, but the significances were different depending on the subjects after 800 seconds.
Estimation of circadian rest-activity rhythm using data average
When using data averaging, the values of amplitude, MESOR, and acrophase were estimated (Figure 2B). Like the results by method of data summation, the estimated values showed a small variance within about 6,000 seconds, but the variance was increased gradually according to the increase of data processing time. Furthermore, the F and p values in zero-amplitude test were statistically significant within about 800 seconds like the results by the method of data summation.
Estimation of circadian rest-activity rhythm using data moving average
When using data moving average, the estimates of amplitude, MESOR, and acrophase were almost constant values within the ranges of all data processing time (Figure 2C). The F values gradually increased according to the increase of data processing time, and the p-values in zero-amplitude test were significant in all ranges from 1 to 21,600 seconds.
Comparison of circadian rest-activity indices using the methods of data average and data moving average
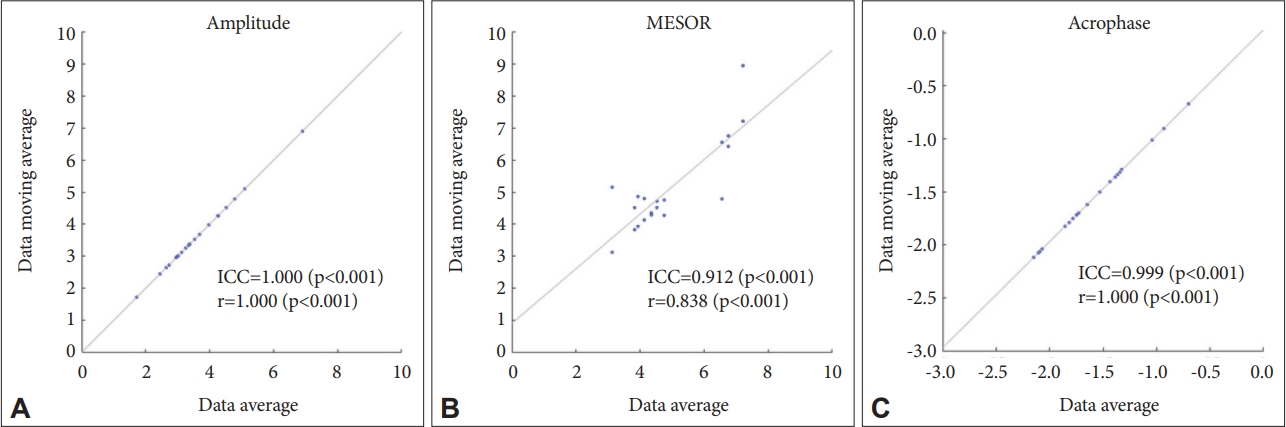
The values of circadian rest-activity rhythm estimated by cosinor analysis using data average and data moving average at the 800-second data processing were compared (Figure 3). These estimated values were compared by ICC and Pearson’s correlation. ICC analysis significantly showed a high concordance rate between two values estimated by data average and data moving average (800-second data processing: amplitude ICC=1.000 [p<0.001], MESOR ICC=0.912 [p<0.001], acrophase ICC=0.999 [p<0.001]). Additionally, there were highly significant correlations between two values estimated by data average and data moving average (800-second data processing: amplitude r=1.000 [p<0.001], MESOR r=0.838 [p<0.001], acrophase r=1.000 [p<0.001]).
DISCUSSION
This study performed mathematical simulations to find the optimal methods of cosinor analysis for studying circadian rest-activity rhythms. As shown in Figure 2C, the method of data moving average showed well-fitted cosine curves independent of the time blocks. Additionally, the estimated circadian indices were constantly maintained in all ranges of time blocks. As the data processing time with data moving average increases, the statistical power was increased. Given that the data moving average reflects the trend of data according to time, the increase of statistical power according to the increase of data processing time can be sufficiently expected. These findings suggest that the cosinor analysis by using data moving average would be an optimal method that could estimate the constant results for an analysis of circadian rest-activity rhythm regardless of data processing time.
In contrast, when using the data summation method, the amplitude or MESOR was inevitably increased as the time block increases as shown in Figure 2A. For this reason, when the circadian parameters were estimated by using data summation, the values of amplitude or MESOR analysis except acrophase cannot be absolutely compared with those by different time blocks. Meanwhile, when using one of data averages, amplitude, MESOR, and acrophase were estimated properly within 800 seconds as shown in Figure 2B. However, as the time block elongates, the variation of the estimated value tended to increase. Locomotor activities can reflect the ultradian rhythms in each mouse from several minutes to several hours except circadian rhythms [26,27]. These ultradian rhythms which might be different in each mouse according to their dopaminergic states can hinder or distort fitting cosine curve around 24 hours [28]. In this study, some results of cosine fitting after about 800 seconds did not show statistical significance. These different results of cosinor analysis might be attributed to individual ultradian rhythms of mice as previously noted. Thus, in order to avoid the influence of ultradian rhythms and properly estimate parameters of circadian rest-activity rhythm, a shorter time block within 800 seconds may be desirable. In addition, this study showed high concordance rate and high correlation between two values estimated by using data average and data moving average with 800-second time block. If a researcher wants to use the method of data average, the optimal time block for analysis would be the range within 800 seconds.
There are some limitations in this study. Firstly, this study only simulated the cosinor analysis for analyzing circadian rest-activity rhythm. Even though cosinor analysis is widely used for circadian rhythm analysis, other analyzing methods for circadian rest-activity rhythm can be used. Thus, these optimal methods on data processing can be adapted when using cosinor analysis. Moreover, this study tested three common methods such as data summation, data average, and data moving average, therefore it is possible that there could be a more optimal method. Furthermore, the analyses in this study were only performed based on the locomotor activities that an optimal strategy for data processing with other circadian indices such as temperature, melatonin, or cortisol should be performed in the future.
Nevertheless, this study suggests the optimal method for analysis of circadian rest-activity rhythm using cosinor analysis. This method can be effectively utilized to analyze circadian rest-activity rhythm using daily locomotor activities in mice. Additionally, when comparing the values estimated by different methods like data average and data moving average, these findings from this study can be helpful to integrate the results of studies using various data processing methods.
Locomotor activity is one of the important characteristics that reflects circadian rhythms. Through using cosinor analysis, the circadian indices such as amplitude, MESOR, or acrophase can be effectively estimated. These simulations suggested that data moving average would be an optimal method for data processing of cosinor analysis. Also, this study showed that the data average within 800 seconds of data processing time might be adaptable. These results can serve as the basic information for research of circadian rhythm using locomotor activities.