INTRODUCTION
The neuropathology of behavioral, metabolic and psychiatric conditions is not sufficiently understood but current knowledge indicates that the underlying mechanisms are, at least in part, overlapping [1,2]. Modification of feeding behavior and energy metabolism are common symptoms of psychiatric disorders, as well as frequent side effects of psychopharmacological treatments [3,4]. The arcuate nucleus of the hypothalamus integrates hormonal, neuronal, and environmental signals to regulate food intake and body weight, but also emotions [1]. Meanwhile, the brain energy requirement at resting state for neuronal computation, information processing and housekeeping functions correspond to more than 20% of the body’s consumption [4-6].
Adenosine monophosphate (AMP)-activated protein kinase (AMPK) is the cellular energy sensor that regulates multiple pathways to maintain energy homeostasis [7]. AMPK is activated under conditions of low intracellular adenosine triphosphate (ATP). The activity of AMPK can be regulated by both allosteric activation and phosphorylation. Allosteric activation is triggered by increased intracellular AMP:ATP or adenosine diphosphate (ADP):ATP ratio, which facilitates the binding of AMP or ADP to the γ regulatory subunit [8]. AMPK phosphorylation at threonine 172 (Thr172) in the α catalytic subunit is mostly regulated by the liver kinase B1 (LKB1) 9 and calcium/calmodulin-dependent kinase kinase β (CaMKKβ) [10,11]. In mammals, metabolism and energy balance at the whole-body level is met by AMPK regulation in the hypothalamus, being activated by fasting and inhibited by feeding and hypothermia [8].
At cellular level activation of AMPK changes metabolism to promote ATP production while switching off biosynthetic pathways that consume ATP [8]. These changes affect not only metabolism of glucose, lipids and proteins, but also mitochondrial biogenesis and autophagy. One target of AMPK is the regulation of acetyl-coenzyme A (CoA) carboxylase (ACC) and carnitine palmitoyl transferase 1 (CPT1) pathway, which is implicated in fatty acid biosynthesis and oxidation. ACC catalyzes the conversion of acetyl-CoA to malonyl-CoA that is a substrate for fatty acid biosynthesis and an allosteric inhibitor of CPT1 [12]. Inhibition of ACC activity by phosphorylation at Ser79 by AMPK [13] results in decreased levels of malonyl-CoA and activation of CPT1 [14]. CPT1 catalyzes the ratelimiting reaction of long-chain fatty acyl-CoA transport to the mitochondria for β-oxidation [15].
Psychoactive drugs frequently influence feeding behavior and body weight [3]. Weight gain in patients treated with clozapine is significantly higher than in patients treated with other antipsychotics [16]. Kim et al. [17] showed that clozapine affects lipid metabolism in rat frontal cortex by activation of the AMPK-ACC-CPT1 pathway. In addition, another study showed that the antipsychotics clozapine and aripiprazole impaired insulin action and increased AMPK phosphorylation in rat hypothalamic neurons [18]. Therefore, we asked what could be the effect of atomoxetine and fluoxetine, two drugs associated with loss of appetite and weight, in the regulation of AMPK phosphorylation and the AMPK-ACC-CPT1 pathway.
Atomoxetine is a selective norepinephrine transporter antagonist [19], which binds to the norepinephrine uptake site with high affinity and the serotonergic uptake site with lower affinity [20]. Atomoxetine is prescribed for attention deficit hyperactivity disorder and is associated with appetite suppression and weight loss [21]. Fluoxetine is a selective serotonin reuptake inhibitor (SSRI), which inhibits reuptake of serotonin through inhibition of the presynaptic membrane serotonin transporter [22]. Fluoxetine is prescribed for depression and anxiety disorders. Fluoxetine is also associated with weight loss and is even considered a potential therapeutic agent for treatment of obesity [23-25]. Here, we investigated the effect of atomoxetine and fluoxetine in the activity of AMPK and downstream ACC and CPT1 pathway in human brain cell lines, SH-SY5Y and U-87 MG cells.
METHODS
Cell culture
Human neuroblastoma cell line SH-SY5Y and human glioblastoma cell line U-87 MG were purchased from the American Type Culture Collection (Rockville, MD, USA). Cells were maintained in Dulbecco’s Modified Eagle’s Medium (Welgene; Seoul, Korea) supplemented with 10% fetal bovine serum (Welgene), 100 U/mL penicillin, and 0.1 mg/mL streptomycin (Welgene) in 95% air and 5% CO2 at 37°C. Cells were seeded at a density of 2×106/mL in 6-well plates or 150 mm dishes. The next day, cell media was changed to serum-free media and cells were treated with different concentration of atomoxetine hydrochloride or fluoxetine hydrochloride (Sigma-Aldrich; St. Louis, MO, USA) dissolved in dimethyl sulfoxide (DMSO). STO-609, a CaMKKα and CaMKKβ inhibitor [26], was purchased from Tocris (Ellisville, MO, USA) was prepared in DMSO and 2.5 μg/mL concentration administered 30 min before atomoxetine and fluoxetine treatments.
Western blotting
Cells were lysed in lysis buffer (2% SDS, 1% 2-mercaptoethanol, 10% glycerol, and 0.1 mg/mL bromophenol blue in Tris-HCl, pH 6.8), heated at 100°C for 10 min and protein amount measured using bicinchoninic acid assay (Bio-Rad; Hercules, CA, USA). 2-mercaptoethanol was added to the lysates (30 μg), heated at 100°C for 5 min and electrophoresed using sodium dodecyl sulphate-polyacrylamide gels. Proteins were transferred to nitrocellulose membranes and incubated with primary antibodies anti-phospho-AMPK (pAMPK [Thr172]; Cell Signaling Technology, Beverly, MA, USA), anti-AMPK, anti-phospho-ACC1/2 (pACC1/2 [Ser79]; Cell Signaling Technology), anti-ACC1/2, anti-β actin antibody (Cell Signaling Technology), anti-CPT1A, and anti-CPT1C (Abcam; Cambridge, UK) for 16 h at 4°C. After washing with Tris-buffered saline-0.05% Tween 20 (TBS-T), blots were incubated with horseradish peroxidase-conjugated anti-rabbit or anti-mouse IgG, and the signal was visualized using the ECL system (Thermo Fisher Scientific; Waltham, MA, USA). Images were acquired using a Molecular Imager ChemiDoc XRS + (Bio-Rad), and band intensity was quantified using Image Lab software version 2.0.1 (Bio-Rad).
CPT1 activity assay
For CPT1 activity assay, mitochondrial and cytosol fractions were isolated from SH-SY5Y and U-87 MG cells using the Mitochondria fractionation kit (Pierce Biotechnology; Rockford, lL, USA). A CPT1 activity was determined using the human CPT1 ELISA Kit (MBS724213, MyBiosource; San Diego, CA, USA) according to manufacturer’s instructions.
RESULTS
Effect of atomoxetine and fluoxetine on the phosphorylation of AMPK and ACC
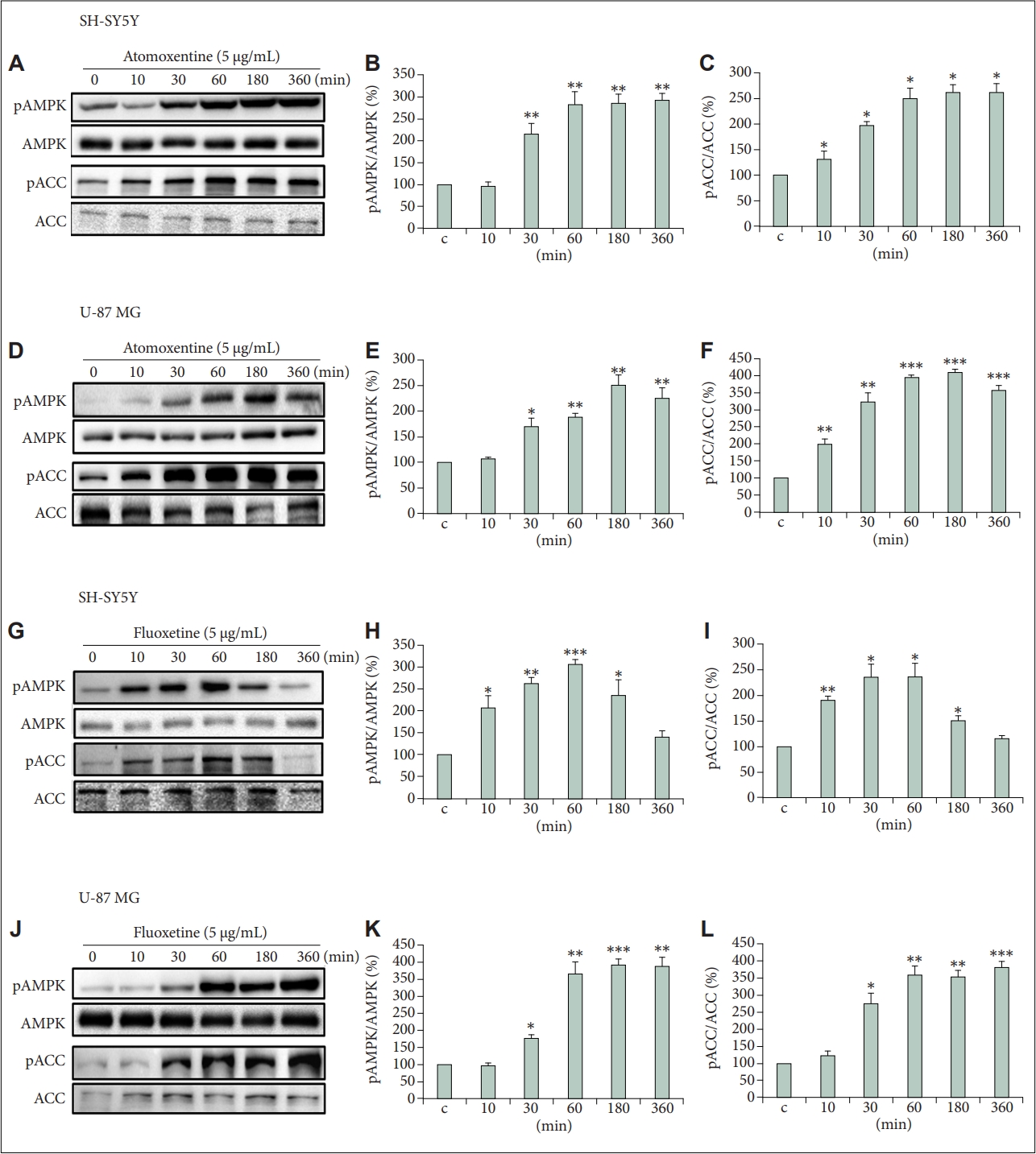
To investigate the effect of atomoxetine and fluoxetine in the activity of AMPK, we treated SH-SY5Y and U-87 MG cells and quantified phosphorylation changes over time (10, 30, 60, 180, and 360 min) using immunoblotting technique (Figure 1). The antibody pAMPK recognizes phosphorylation at Thr172 sites, which is directly correlated with AMPK activity [27]. Overall, phosphorylation of AMPK increased significantly by at least 2-fold in the two cell lines with atomoxetine and fluoxetine treatment (Figure 1A, B, D, E, G, H, J, K). However, although the response kinetics to atomoxetine was similar in the two cell lines, the response to fluoxetine was different. The expression levels of pAMPK in SH-SY5Y cells treated with fluoxetine followed a bell-shaped curve with an early increase after 10 min and decreasing after 60 min (Figure 1H). For the other conditions, a significant increase in AMPK phosphorylation was observed 30 min after drug treatment with a maximal constant level after 180 min (Figure 1A, E, K). As expected for an AMPK substrate, the changes in the phosphorylation levels of ACC1/2 (or ACC for simplicity) followed closely those observed for AMPK (Figure 1A, C, D, F, G, I, J, L). Therefore, these results suggest that atomoxetine and fluoxetine activate AMPK and inactivate ACC, a downstream target on the AMPK-ACC-CPT1 pathway.
Mitochondrial CPT1 activity is increased by atomoxetine and fluoxetine
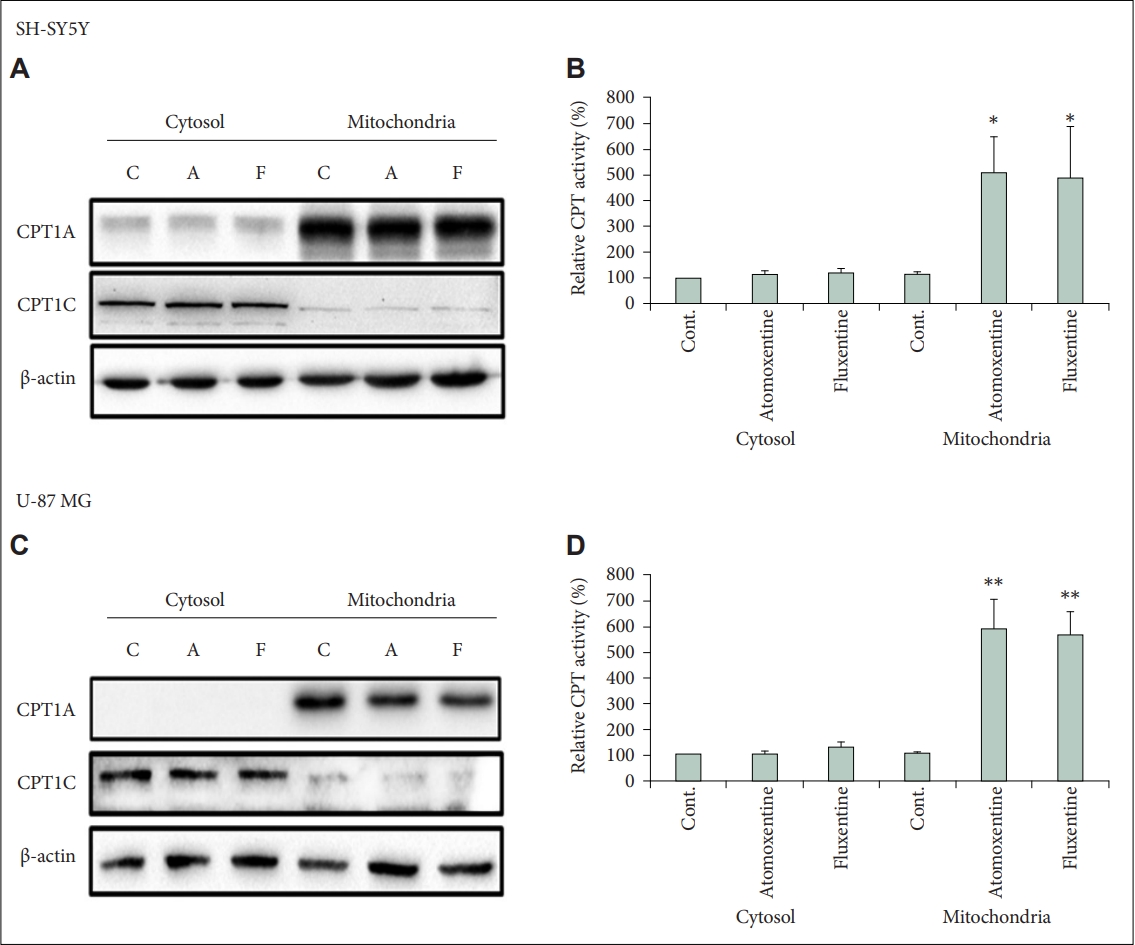
The increase in ACC phosphorylation suggests that CPT1 expression may be increased after atomoxetine and fluoxetine treatment. Two CPT1 isoforms are present in the brain; mitochondrial CPT1A is expressed throughout the body and CPT1C is localized in the endoplasmic reticulum and expressed in neurons [28]. The exact function of CPT1C is not completely understood but it is supposed to facilitate the entry of palmitoyl-CoA to the endoplasmic reticulum [28]. Mitochondrial CPT1 control fatty acid β-oxidation by catalyzing the first step in the transport of long chain fatty acids from the cytoplasm to the mitochondrial matrix. 29 Using immunoblotting we observed that both cell lines express CPT1A and CPT1C, but protein levels did not change with the two drug treatments (Figure 2A and C). Therefore, we measured CPT1 activity in mitochondrial and cytosol cellular fractions after atomoxetine or fluoxetine treatment (Figure 2B and D). Only mitochondrial fractions show modified CPT1 activity with drug treatment. For atomoxetine treatment, activity was increased 5.1-fold in SH-SY5Y and 5.9-fold in U-87 MG cell lines. For fluoxetine treatment, activity was increased 5.0-fold in SH-SY5Y and 5.7-fold in U-87 MG cell lines. These results suggest that atomoxetine and fluoxetine treatments increase fatty acid β-oxidation in the mitochondria.
Atomoxetine activate AMPK through CaMKK signaling pathway
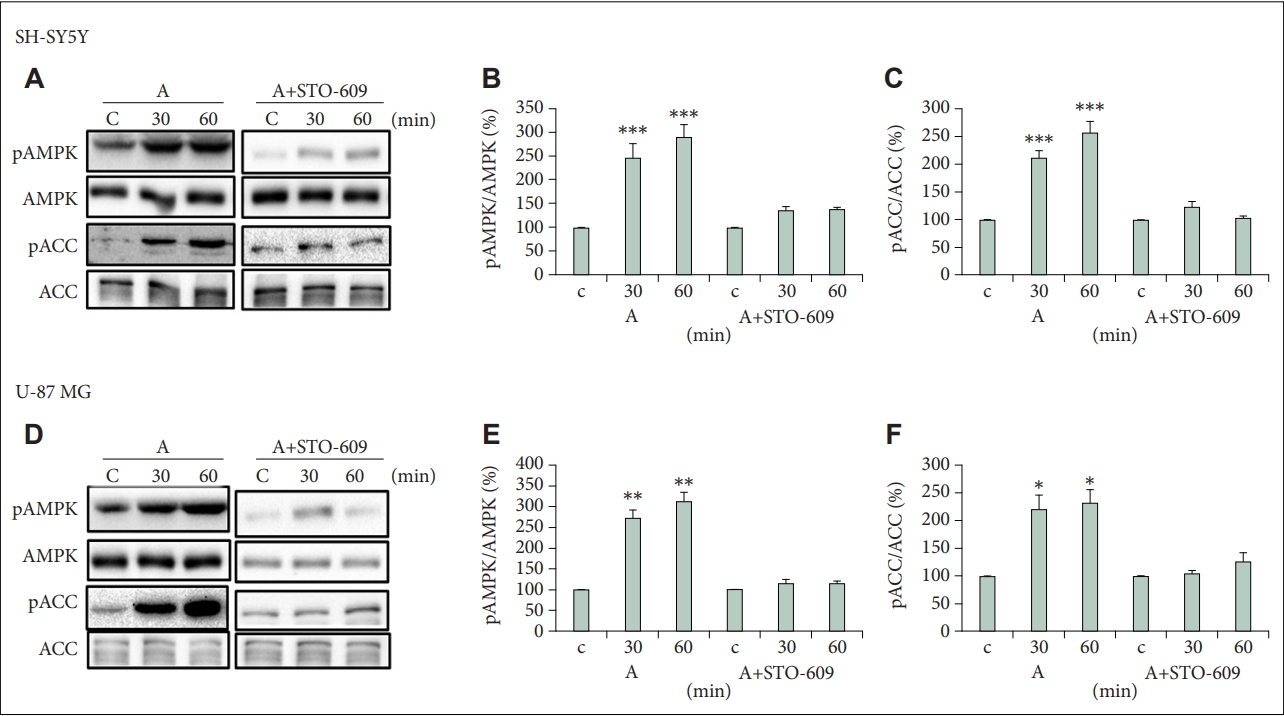
AMPK is activated by CaMKK [11], therefore we tested if the activation effect of atomoxetine on AMPK could be mediated by CaMKK. For this we used STO-609, which specifically inhibits both CaMKKα and CaMKKβ isoforms, and tested AMPK and ACC phosphorylation after 30 and 60 min atomoxetine treatment that correspond to maximal expression of pAMPK and pACC (Figure 2). Induction of pAMPK expression by atomoxetine as well as pACC expression were dramatically decreased by treatment with STO-609 in SH-SY5Y and U-87 MG cells (Figure 3). In SH-SY5Y cells the expression levels were decreased from 2.9- to 1.4-fold for pAMPK and from 2.6- to 1.0-fold for pAAC after 60 min treatment (Figure 3A-C). In U-87 MG cells the expression levels were decreased from 3.1- to 1.2-fold for pAMPK and from 2.3- to 1.3-fold for pAAC after 60 min treatment (Figure 3D-F). These results show that atomoxetine activate the AMPK-ACC-CPT1 pathway by means of CaMKKβ activity.
DISCUSSION
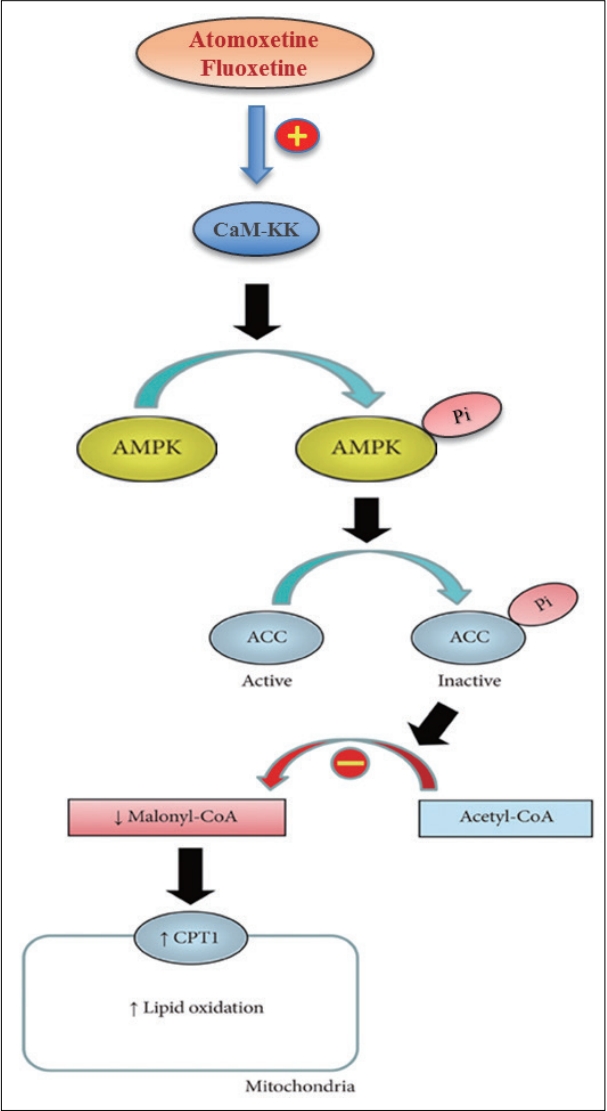
In this study, we investigated the effect of atomoxetine and fluoxetine, two drugs that are associated with loss of appetite and weight, on the regulation of AMPK-ACC-CPT1 pathway in human brain cell lines (SH-SY5Y and U-87 MG cells). We show that atomoxetine and fluoxetine induce the AMPK-ACC-CPT1 pathway (Figure 4). The specific increase in activity of mitochondrial CPT1 induced by atomoxetine and fluoxetine suggests that these drugs induce mobilization of intracellular lipid stores to generate ATP from fatty acid β-oxidation in SH-SY5Y and U-87 MG cells. These findings seem contradictory to those observed in the frontal cortex of rats treated with a clinically relevant dose of clozapine, an antipsychotic agent associated with heavy weight gain, where activation of AMPK-ACC-CPT1 pathway was also observed [17]. However, clozapine induces the isoform CPT1C and not the CPT1A [17], which is the opposite of what is observed in this study with atomoxetine and fluoxetine treatment. CPT1C proposed role is to facilitate palmitoyl-CoA entry into the endoplasmic reticulum [28], therefore it is possible that clozapine affects lipid composition of neuronal membranes in the frontal cortex [17]. Another reason for the discrepancy from data from the frontal cortex of rats treated with clozapine may be the use of SHSY5Y and U-87 MG lines, which are neuroblastoma and glioblastoma cells in this study. The energy metabolism of cancer cell may differ from normal neurons [30].
Our results highlight the complexity of appetite regulation by AMPK, which activity is hormone and tissue specific [31]. For example, leptin activates AMPK in skeletal muscle whereas it has the opposite effect in the hypothalamus [32,33]. Nevertheless, numerous studies in rat hypothalamus show that appetite is correlated with AMPK-ACC-CPT1 pathway activity. Malonyl-CoA is markedly increased by feeding and suppressed by fasting [34]. In agreement with this, a decrease in malonyl-CoA and consequent CPT1 activation in the mediobasal hypothalamus of rats resulted in increased food intake and obesity [35]. Conversely, genetic or biochemical inhibition of hypothalamic CPT1 activity decreased food intake [36]. In order to better understand the effect of atomoxetine, fluoxetine, and clozapine in patient feeding and weight gain and in cellular AMPK activity it would be interesting to analyze the hypothalamus, as well as peripheral organs, in treated animals.
Previous studies have shown that various areas of neurotransmission altered by antipsychotics may affect energy and glucose regulation [37] and that antipsychotics with the most intense metabolic effect are the most effective agents [38]. These finding suggests that metabolic effects and clinical efficacy of psychiatric medication may not be an independent phenomenon but rather intricately related [4]. The downstream pathways of AMPK involve the mammalian target of rapamycin (mTOR) pathway, cell cycle regulation and autophagy (a lysosome-dependent self-degradation process of cellular components). These downstream pathways are implicated in neuronal plasticity which most likely play a major part in the therapeutic effects of antipsychotic medication [39,40]. Therefore, AMPK as a metabolic sensor and a key regulator of cellular homeostasis may be an important player in the interaction between metabolic effect and clinical efficacy of psychiatric medication.
Atomoxetine induce the AMPK-ACC-CPT1 pathway through activity of CaMKKβ in SH-SY5Y and U-87 MG cells. LKB1 and CaMKKβ are two upstream kinases that phosphorylate Thr172 of AMPKα subunit [11,41]. LKB1-dependent AMPKα phosphorylation at Thr172 is enhanced by the binding of AMP to the AMPK γ subunit [42]. CaMKKβ activates AMPK in response to increases in cellular calcium without significant change in ATP/ADP/AMP ratio [43]. CaMKKβ is expressed primarily in the brain and therefore the calcium mediated pathway may be the most relevant in neurons [44]. Previously, it was reported that knockdown of CaMKKβ using siRNA increased cell death following oxygen-glucose deprivation (OGD), whereas inhibition of CaMKKβ by STO-609 significantly and selectively down-regulated levels of phosphorylated SIRT1 after OGD [45]. Therefore, AMPK pathway activation by CaMKKβ was confirmed using STO-609.
Our results show that atomoxetine and fluoxetine-activated AMPK increase the phosphorylation of ACC in human SH-SY5Y and U-87 MG cells. Next, we showed that the activation of ACC leads to the increase in fatty acid beta oxidation in mitochondria. Using STO-609, we showed that AMPK-ACC-CPT1 pathway was activated by CaMKKβ. In conclusion, we show that atomoxetine and fluoxetine treatments may increase fatty acid β-oxidation in the mitochondria through activation of AMPK-ACC-CPT1 pathway in human SH-SY5Y and U-87 MG cells, thus mimicking fasting since AMPK is activated but not necessarily accompanied by increase in appetite as energy is produced. One limitation of this study is the use of SH-SY5Y and U-87 MG lines, which are neuroblastoma and glioblastoma cells. Future studies are necessary to explore the activation of AMPK by atomoxetine and fluoxetine after differentiation of SH-SY5Y cells into neurons, along with in vivo systemic metabolic effects on multiple organs, such as liver, adipose tissue, muscle, and the brain. Comparison of fatty acid oxidation in mitochondria among different anti-psychotics are also warranted in the future.