INTRODUCTION
Humans live their daily lives adapting to the 24-hour lightdark cycle produced by the Earth’s rotation. Light gives humans the most potent cues for synchronizing their circadian rhythms, and plays a major role as an entraining environmental cue for maintaining those rhythms. Light exposure at inappropriate times can disturb human circadian rhythms, and is known to have a profound effect on physical and mental health [1]. Jet lag and shift work are common causes of inappropriate light exposure and circadian misalignment, resulting in various problems [2,3]. Moreover, seasonal variations in solar radiation are reported to be closely related to suicide rate and aggravation of mood disorders [4-6], suggesting that humans are sensitive to light exposure, whether at night or day. Thanks to the invention and development of electric lighting, our lives were made more productive and leisurely by proper and effective use of light at night (LAN). However, there are reasons to believe that the influence of LAN is not entirely positive [7], because even as it has become a universal feature of modern society, various negative influences of LAN have emerged [8].
In addition to social, ecological, and behavioral consequences, exposure to LAN may have negative impacts on sleep and circadian rhythms, resulting in poor sleep and circadian misalignment [7]. Because exposure to LAN is so common, we tend to overlook its influences on our health, and habitually prolong our exposure to it. As reported by several epidemiological studies, long-term and repeated exposure to LAN might have a negative effect on human health, resulting in an increased risk of physical and mental disorders [9-12]. Melatonin, which is closely related to human sleep, has been reported to be very sensitively suppressed by light exposure. Due to the sensitivity of melatonin to light exposure, melatonin has been regarded as an important hormone in explaining the mechanism of the effects of LAN exposure on humans [13].
Studies of the effects of LAN exposure on humans have mostly used bright light, while relatively few studied the effects of dim LAN (dLAN) exposure. This is probably due to the assumption that the effect of dim LAN on humans is not as great as bright LAN, and the study design of the impact of dLAN on human is relatively challenging. But we cannot ignore that exposure to dLAN is a common problem, as a consequence of light pollution from smart devices, television, and street lights shining through windows. In both animal and human studies, exposure to dLAN has been shown to have impacts on metabolism [14,15], immune function [16], depressive or anxiety-like phenotype [17,18], and the circadian system [19]. We previously performed an experimental study to investigate the effect of dLAN exposure on sleep in healthy young male subjects, and reported an increase in awakening and REM sleep as a consequence of exposure to dLAN, as assessed by nocturnal polysomnography (NPSG) [20], and dLAN during sleep was shown to have an effect on some aspects of fatigue [21]. Since dLAN become a major environmental issue in many countries, exposure to dLAN is regulated by light intensity limits of 10 lux or less (such as 5 lux) [22]. Although there have been several studies reporting the negative effect of dLAN exposure on humans, they did not clearly demonstrate mechanisms that could explain the effects on sleep, but suggested that changes in melatonin secretion could be one of the main factors [23,24].
In this study, we investigated the impact of dLAN exposure during sleep in healthy young female subjects with an experimental study using the same design as our previous study of male subjects [20]. To investigate the difference in sleep changes between 10 lux and 5 lux, which is a weaker intensity, the study sample was divided into the two dLAN groups. In addition, salivary melatonin was measured both before sleep onset (the beginning of time in bed) and after sleep offset (the end of time in bed), to find out indirectly whether melatonin changes occur under dLAN exposure. Through direct and indirect experimental investigation, we aimed to identify not only the effects of dLAN exposure on sleep in young women, but also their association with changes in salivary melatonin level. We also analyzed the impacts of dLAN during sleep in total (female and male) subjects, and compared the differences according to gender.
METHODS
Participants
Between September 2014 and March 2015, a total of 28 young female subjects ranging from 19 to 35 years of age (mean±SD: 23.86±2.97) were recruited for this study through online advertising, within the intranet of Korea University students, to find volunteers who “sleep like a baby.” Prospective subjects were screened regarding their sleeping conditions and physical and mental health, through questionnaires including the Mini International Neuropsychiatric Interview [25] and face-to-face interviews with two clinicians (HJL and CHC) who specialize in sleep. After the initial screening, we randomly divided the participants into two groups according to the dLAN intensity to which they would be exposed (Group A: 5 lux, Group B: 10 lux). They were blinded to the group assignment. To investigate the impact of dLAN during sleep in a larger population sample, we collected female subjects from this study and male subjects from a previous study [20] that used the same design and protocol (mean age±SD of total sample: 23.04±2.92).
During the entire study schedule (from one week prior to the experiment until its end), the subjects were instructed to maintain their usual sleep-wake cycles, and not to use medicine, caffeine, or alcohol, which could affect their sleep. One week prior to the experiment, an actigraph (Actiwatch-L, Mini Mitter) was provided, to be worn for the purpose of checking whether they maintained regular sleep-wake cycles. We excluded participants who showed sleep efficiency (SE) below 90%, or an apnea-hypopnea index (AHI) >5, in the first NPSG session (Night 0). As a result, 3 subjects were excluded from the study on the basis of SE values below 90%. The remaining 25 participants served as subjects for the study (12 for Group A and 13 for Group B). With the 23 male subjects from the previous study for the comparison according to gender, and a total of 48 healthy young subjects (female and male) were included (23 for Group A and 25 for Group B). All participants provided informed written consent for the study, which was approved by the Institutional Review Board (IRB) of Korea University Anam Hospital (IRB No. ED12261). All aspects of the study were conducted in accordance with the Declaration of Helsinki.
Protocol
Beginning one week prior to the experiment, we instructed the subjects not to nap, and requested that they maintain regular nighttime sleep cycles, which were checked by actigraph data. The experimental study was performed over three consecutive nights. For every NPSG session, the time in bed was controlled from 11 P.M. to 7 A.M., and on Nights 1 and 2 staff obtained salivary samples at two time points, just before sleep onset and just after sleep offset, using only dim red light under 5 lux so as not to influence melatonin levels by light exposure. To minimize first-night effects, we performed an NPSG session with no light on the first night (Night 0) of the study. The main experiment started on the second night (Night 1). The participants underwent an NPSG session with no light during sleep, the same experimental conditions as on Night 0, except that salivary melatonin samples were collected. On the following night (Night 2), each participant underwent an NPSG session, with dLAN according to the randomly assigned group (5 or 10 lux) during the entire sleep, but blinded with respect to whether they were in the two different light groups. A light box was installed in the sleep laboratory on the upper part of the wall opposite the participant’s head, consisting of light emitting diodes (LED) as the light source, with a broad spectrum (peak λ: 463.6 nm, center λ: 467.6 nm, centroid λ: 554.3 nm, dom λ: 501.4 nm, Correlated Color Temperature: 5779.1 K, General Color Rendering index: 90). To ensure precision, staff checked the luminous intensity at the participant’s eye level in a supine position, using an illuminometer (ANA-F11, Tokyo photo, Tokyo, Japan). More detailed information on the study protocol, excluding melatonin measurements, can be found in our previous report [20].
Measurements
Sleep states were recorded by NPSG on each experimental night. Data from electroencephalograms (EEG), electrooculograms (EOG), and chin electromyograms (EMG) were assessed to identify sleep stages, arousals, and movements. Respiratory events were identified using data from abdominal and chest bands, a pulse oximeter, a position sensor, and airflow monitoring devices (nasal pressure plus thermistor). Sleep states were scored according to standard criteria, by a well-trained PSG technician. The measured variables included total sleep time (TST), sleep efficiency (SE), wake time after sleep onset (WASO), sleep latency (SL), Stage N1 time, Stage N2 time, Stage N3 time, Stage R time, Stage R latency, REM density, apnea-hypopnea index (AHI), respiratory effort-related arousal index (RERA index), respiratory disturbance index (RDI; AHI+RERA index), percentage of supine position (Supine), periodic limb movement during sleep index (PLMS index), limb movement index (LM index), total arousal time (TA), and spontaneous arousal time (SA). The time for each sleep stage was converted to the corresponding fraction of TST (percentage) for analysis.
Salivary melatonin was measured by well-trained staff just before and after sleep in the NPSG room, under extremely dim red light (less than 5 lux) to prevent any confounding effect on melatonin secretion. The saliva samples were placed in dark brown e-tubes, 0.5 mL each, to avoid any unwanted effects of light, and stored at -80°C immediately after sampling. The samples were analyzed within 24 hours of collection, by an enzyme-linked immunosorbent assay and an enzyme-linked immunospecific assay, using a Melatonin ELISA Kit (Saliva, Salimetrics, Carlsbad, CA, USA) according to the manufacturer’s instructions.
Statistical analysis
Demographic and sleep characteristics, as well as salivary melatonin level before sleep on Night 1, were compared using Student’s t-test or the Mann-Whitney U test. To investigate melatonin level changes, we compared the salivary melatonin levels measured before sleep onset and after sleep offset, calculating, 1) the area under the line connecting the two melatonin level points for each night, 2) the melatonin level after sleep offset minus the level before sleep onset, and 3) the melatonin level after sleep offset. Using repeated measures analysis of variance (ANOVA) on the NPSG-derived sleep variables in the female and merged subjects, and the 3 melatonin parameters from Nights 1 and 2 in the female subjects, we analyzed the effect of dLAN on each value across groups and nights. Values of p<0.05 were considered statistically significant. All statistical tests were performed using SPSS for Windows ver. 16 (SPSS Inc., Chicago, IL, USA).
RESULTS
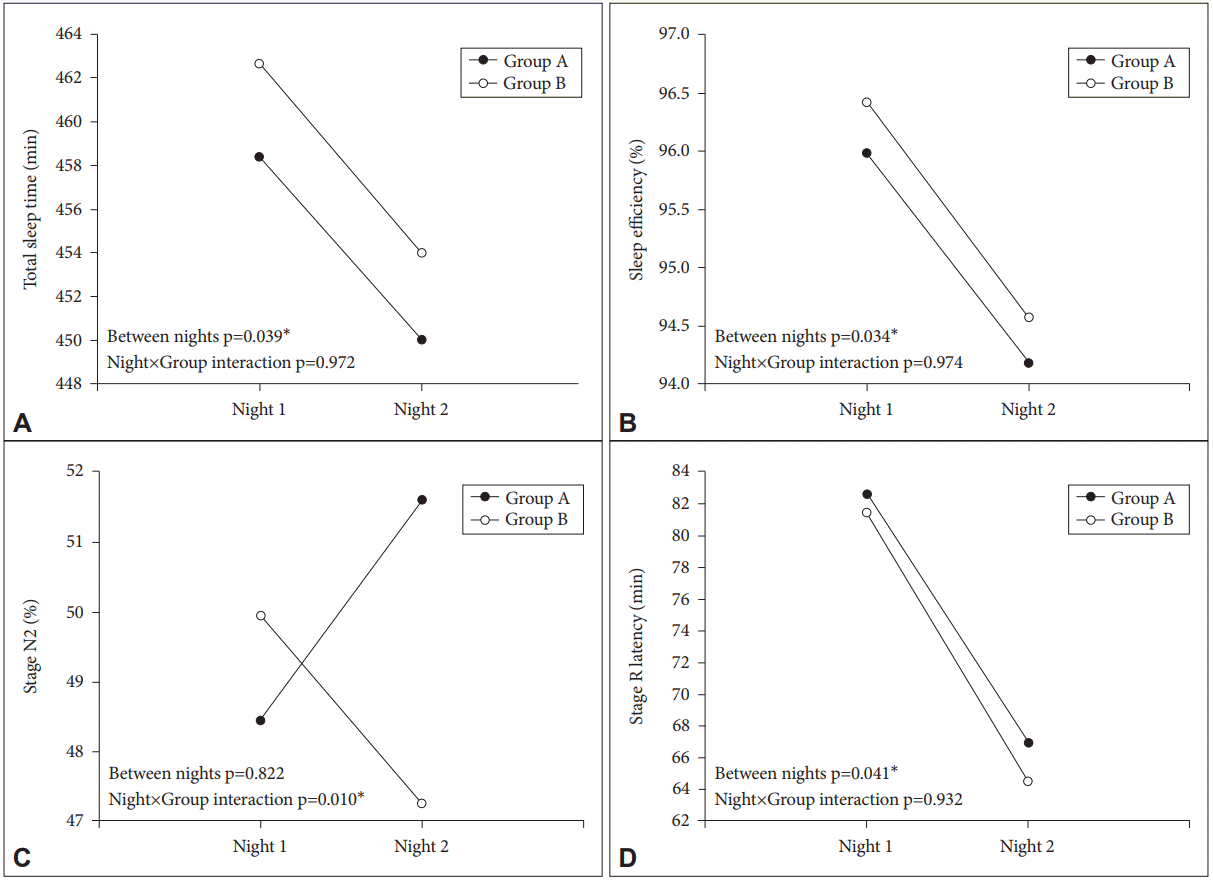
First, we investigated the effect of exposure to dLAN on sleep and melatonin in healthy young female subjects. Table 1 shows the results of repeated measures ANOVA on NPSG-derived sleep variables across nights and groups, in female subjects. We could find statistical significance for decreased TST (F=4.818, p=0.039), SE (F=5.072, p=0.034), and Stage R latency (F=4.664, p=0.041) between nights, but no significant nightgroup interactions. Stage N2 showed a significant night-group interaction (F=7.879, p=0.010), but no significant difference between nights. Figure 1 shows the significant findings for sleep variables in the female subjects.
Table 2 shows the results of a repeated measures ANOVA on melatonin changes across nights and groups, in the female subjects. For the 3 variables indirectly reflecting melatonin levels (area under the curve; difference between level before and after sleep; level after sleep), we did not find any statistically significant effect of night, or any night-group interaction. These results suggest that exposure to dLAN did not significantly influence melatonin secretion.
Variations in NPSG-derived sleep variables across nights and groups were investigated in the total (male and female) sample. As shown in Table 3, we found statistical significance for decreases in TST (F=14.971, p<0.001) and SE (F=7.687, p=0.008), and increases in WASO (F=6.322, p=0.015) and Stage R time (F=5.031, p=0.03). Significant night-group interactions were found for Stage N2 time (F=7.590, p=0.008) and Stage R time (F=4.579, p=0.038).
Lastly, comparison of sleep variables across nights according gender in 5 or 10 lux experimental condition was investigated, as shown in Table 4. When comparing within 5 lux experimental condition, we could find statistical significance for decrease in TST (F=5.802, p=0.025) between nights in both gender group. Significant night-gender group interactions were found for Stage N2 time (F=4.376, p=0.049) and Stage R latency (F=5.909, p=0.024). Within 10 lux experimental condition, we found statistical significance for decreased TST (F=10.48, p=0.004), SE (F=5.409, p=0.029), Stage N2 time (F=16.52, p<0.001), and Stage R latency (F=5.575, p=0.027) and for increased WASO (F=7.297, p=0.013) and Stage R time (F=8.482, p=0.008) between nights in both gender group.
DISCUSSION
From this study of healthy young female subjects, we obtained several results concerning the impact of dLAN exposure on sleep. Total sleep time and sleep efficiency were significantly decreased on Night 2 compared with Night 1, suggesting that dim light exposure during sleep may impact negatively on sleep quality and quantity. We previously performed an experimental study to examine the impact of dLAN on sleep in healthy young male subjects, and reported changes in sleep, including decreased Stage N3 and increased wake time after sleep onset, after dLAN exposure during sleep, proposing that dim light exerts a negative effect on quality of sleep by promoting awakening and decreasing deep sleep [20]. In the analysis of the merged male and female subjects, the effect of dLAN exposure on human sleep becomes clear. TST and SE were significantly decreased, and WASO and Stage R were increased, on Night 2 compared with Night 1. In this study, we replicated the negative effect of dLAN exposure on the quality and quantity of sleep in female subjects and in the merged group of subjects. This result has similar implications to our previous findings obtained using male subjects, but extends them across sexes.
Other studies have also reported that exposure to dLAN influences the circadian system and sleep. Shuboni and Yan investigated the disturbing effects of dim light exposure on the regulation of circadian rhythms in an animal study, and reported that dLAN exposure can cause changes in circadian function, suggesting this as one of the mechanisms underlying the adverse effects of dLAN [26]. Another animal study, performed by Fonken et al., reported that exposure to dLAN altered core circadian clock rhythms in the hypothalamus at both the gene and protein levels, and was associated with altered feeding behavior and increased weight gain [27]. However, a different animal study reported that chronic dLAN did not significantly affect sleep timing or quality of sleep [28]. Taking all this together, we could conclude that dLAN has a generally negative effect on sleep and circadian system; but human studies have been relatively sparse. Therefore, the result of this study is meaningful in that it was an experimental study performed with human subjects, and that generally similar results were obtained as in previous studies.
A significantly shortened Stage R latency was found in female subjects exposed to dLAN during sleep. Our previous research performed in male subjects found a significant correlation between dLAN exposure and an increase in REM sleep time [20]. Though the same results were not obtained, the findings of shortened Stage R latency and increased Stage R time could be considered in a similar context in terms of an increase in REM sleep. Borniger et al. [28] reported that dLAN-exposed animals showed increased REM sleep at specific time points near the dark/light transition, suggesting an influence of chronic dLAN exposure during sleep on the organization of REM sleep. Several results, including our previous and present studies, showed increased REM or reduced Stage R latency. Interestingly, the effect of dLAN exposure on human REM sleep was more pronounced in the merged male and female sample. In this extended sample, we could confirm that dLAN exposure not only significantly increased REM sleep, but also increased REM sleep more pronouncedly under 10 lux than under 5 lux, that is, there was a night-group interaction. Given only the results of this study, the increase in REM sleep cannot be clinically confirmed as either good or bad. Nonetheless, based on previous studies, an increase in REM sleep due to dLAN exposure could be considered to have a somewhat negative impact on humans. Depression is known to be associated with altered REM sleep parameters, such as shortened REM latency, increased REM duration, and increased REM density [29], changes resembling those seen in this study. Riemann et al. [30] proposed that instability of REM sleep contributes to disrupted sleep and insomnia, based on evidence of increased micro- and macro-arousals during REM sleep in insomnia patients.
When comparing the gender difference of impact of dLAN on sleep, we could not find any notable significant gender difference. Instead, consistent sleep changes were observed in both male and female groups under experimental conditions of 10 lux. TST, SE, Stage N2 time, and Stage R latency were decreased under 10 lux dLAN exposure, and WASO and Stage R time were increased. This result shows that 1) the no gender difference of dLAN exposure on sleep, 2) a significant impact on sleep of the light intensity of 10 lux rather than 5 lux, and 3) the decrease of amount and quality of sleep and the increase of REM under the condition of 10 lux.
Even though Stage N2 or Stage R latency showed a significant night-group interaction in present analysis, this result should be interpreted cautiously, because there was no significant difference between nights, in other words, the values of the two groups crossed over from Night 1 to Night 2. A nightgroup interaction is not easy to interpret unless the two groups both exhibit trends in the same direction. Thus, we refrain from giving too much meaning to the outcome of a significant nightgroup interaction without significant night difference in this study.
To explore the possible role of melatonin changes as one of the causes of sleep changes induced by dLAN exposure, we measured salivary melatonin levels before and after sleep in a controlled experimental setting, and investigated whether significant melatonin changes were caused by dLAN exposure. However, we could not detect any significant changes in melatonin, suggesting no effect of dLAN exposure on melatonin secretion. It has long been known that light-sensitive melatonin is not significantly inhibited by dim light. This fact manifests itself in the dim light melatonin onset (DLMO) measurement, a representative method for measuring melatonin [31]. According to the instructions for the DLMO measurement method, bright light should be blocked from 1 hour before the start of the measurement, and subjects should be kept in a dim light environment of 10 lux or less [32]. In other words, this suggests indirectly that dim light of 10 lux or less does not have a significant effect on the secretion of melatonin. In this study we also found no suppression of peripheral melatonin (saliva) by dLAN exposure. However, the effect of dim light on human melatonin secretion is not yet a settled question. In animal studies, several researchers found that dLAN of 1.08-5 lux inhibited pineal melatonin production and was probably sufficient to suppress melatonin [17,33]. In human studies, the results of animal experiments might not easily be replicated, because it is difficult to conduct direct (invasive) measurements on the pineal gland, which is the key to melatonin secretion.
When the photoreceptive ganglion cells (pRGCs) in the eye detect light exposure, the resulting neural activity is conveyed to neurons that make up the circadian system in the suprachiasmatic nucleus [34]. Melatonin synthesis is largely controlled through this neuronal system, starting with exposure to light as well as direct sympathetic control of physiological processes caused by lighting conditions [35,36]. Because physiological states can be altered through various mechanisms as a consequence of light exposure [7], even though we did not obtain meaningful results in our study, we still cannot resist suggesting a change in melatonin due to dLAN exposure as the strongest cause of sleep changes in this study. Of course, it is necessary to look for other factors besides melatonin to explain the effect of dLAN on human’s sleep. Another factor that might explain this research finding is melanin-concentrating hormone (MCH), as we mentioned in our previous report [20]. Based on reports that the firing of MCH-expressing neurons is very sensitive to light exposure [37], and that stimulation of MCH is associated with increased REM sleep [38,39], the possibility should be considered that dLAN exposure causes changes such as increased REM sleep or shortened Stage R by somehow affecting MCH.
This study has some limitations. First, relatively few subjects were included. For comparability with our previous study, we used the same number of subjects. By performing a controlled experimental protocol, it was possible to overcome this limitation. A repeated measures ANOVA, which was the main statistical method in this study, is a reasonable and appropriate method for analyzing significant differences between two consecutive nights in the same group of subjects. Moreover, we collected the samples for both sexes, to increase the statistical power and draw out meaningful findings. Second, in the measurement of melatonin, direct measurements such as blood sampling and continuous sequential detection were not performed. In this study, salivary melatonin was obtained and measured twice in a night, before and after sleep. Therefore, it was not possible to directly measure the changes in melatonin caused by dLAN, nor to detect the trend of change. It would be challenging to make continuous melatonin measurements, which are invasive and interfere with sleep, in human subjects. Unfortunately, the lack of melatonin measurements should be carefully considered in the interpretation of the study results. Third, we did not use a counterbalanced study design, because we thought that the impact of dLAN on sleep might influence sleep on the following day [40]. Because there are advantages and disadvantages of each research design, no design could possibly be satisfactory in every respect. Fourth, female’s menstrual cycle is known to be associated with sleep and melatonin [41], but this study did not control the menstrual cycle. It was very difficult to control the menstrual cycle in experimental studies on humans, and this can be regarded as a limitation in this study. Future research will need more detailed and sophisticated experimental designs.
In conclusion, from this experimental study performed on healthy young female subjects and merged male and female subjects, we obtained generally consistent results concerning the negative impact of dLAN exposure on the quality and quantity of sleep. People in modern society who are exposed to dLAN during sleep, from sources such as smart devices, television, and street lights shining through the window, could show interference in their sleep, resulting in a decrease in total sleep time and poor sleep efficiency. Whether melatonin changes occur as result of dLAN exposure remains unclear, but this is still the most probable mechanism to explain the negative impact of dLAN on human sleep. Further studies focusing on the basic physiologic processes induced by dim light exposure at night should be performed, to clarify the core mechanisms of dLAN. Considering the frequency of exposure to various light sources at night in our routine lives, we should control or prohibit exposure to dLAN for our health, as well as the soundness of our sleep.