INTRODUCTION
Schizophrenia is a complex neuropsychiatric disorder, affecting about 1% of the general population [1]. This disease is characterized by a wide spectrum of symptoms, which include thought disorder, perceptual aberrations, cognitive difficulties, apathy and social withdrawal. Evidence from family, twin and adoption studies support high heritability in the development of schizophrenia, although the exact causes are unclear, genetic and environmental factors contribute to the etiology of schizophrenia [2].
Schizophrenia has a higher frequency of smoking (over 80%) than the normal population (23%-30%) [3,4]. In addition, case reports showed that psychotic symptoms would relapse when patients stopped smoking [5]. Nicotine has been reported to normalize two psychophysiological dysfunctions in schizophrenia: an auditory sensory gating deficit and abnormal smooth pursuit eye movement [6,7]. Brain PET Imaging study found that the ╬▒7-nAChR was lower in the cingulate cortex, frontal cortex, and hippocampus of Schizophrenia patients [8]. The diminished nAChR is a genetically transmitted trait related to the risk for schizophrenia, rather than the effect of smoking or antipsychotic treatment [9,10]. As well known, typical antipsychotics are thought to exert their effects primarily by antagonizing dopamine D2 receptors leading to the dopamine hypothesis of schizophrenia [11]. Pre-synaptic Nicotinic receptors can mediate the release of dopamine and this points to neuronal nicotinic receptors as important candidate genes in this disorder [12,13].
Nicotine exerts its effect through neuronal nicotinic acetylcholine receptors expressed in the brain and periphery. Some of the nicotinic receptor subunit genes, such as CHRNA7 and CHRNA4 have been investigated the association with schizophrenia or other psychiatric disorders in genetic studies [14]. The ╬▒7 neuronal nicotinic receptor subunit gene (CHRNA7), was first genetically linked to the P50 auditory sensory processing deficit in schizophrenia, and then to schizophrenia as a disease in multiple independent studies [15-17]. Several studies have found decreased expression of the CHRNA7 gene in the postmortem hippocampus, reticular thalamic nucleus and frontal cortex of schizophrenic subjects. The ╬▒7 receptor agonists are ranked as the most promising targets for developing drugs to treat cognitive impairments in the disorder [18]. Recently, some studies indicated that variations in the gene CHRNA4 may modulate attention and spatial scaling performance in humans [19,20]. Besides, CHRNA5 was found to be associated with nicotine dependence and CHRNA3 was found to be associated with prepulse inhibition (PPI) [21,22].
Therefore, to further evaluate the role of the genes above in susceptibility to schizophrenia, we conducted a case-control study in the Han Chinese population. We selected one SNP for CHRNA3, CHRNA4, CHRNA7, and two SNPs for CHRNA5 according to the previous studies and the minor allele frequency of the SNPs was higher than 0.05 [19,23,24]. Meanwhile, we explored the interactive effect among the SNPs we studied using the multifactor dimensionality reduction (MDR) method, which has great advantages versus the conventional statistical approaches for detecting gene-gene interactions of the genes selected. Complex interactions among genes and environmental factors are known to play a role in common human disease etiology. However, the identification and characterization of gene-gene interactions for common complex human diseases remain a challenge for human geneticists. Traditional statistical methods are not well suited for detecting such interactions, especially when the data are highly dimensional (having many attributes or independent variables) or when interactions occur between more than two polymorphisms. To address these issues, a variety of bioinformatics methods for identifying gene-gene interactions have been developed, and one such method is MDR [25], a data mining tool for detecting and characterizing nonadditive interactions between discrete variables that affect binary results.
METHODS
Subject
A total of 1,035 schizophrenic patients (526 males and 509 females) at Shanghai Mental Health Center, Shanghai Jiao Tong University were included in this study (mean age 33.7┬▒9.2 years). Diagnoses were made based on DSM-IV. Patients with other mental disorders and severe physical disorders were excluded. The control group consisted of 816 subjects (420 males and 396 females, mean age 33.3┬▒10.6 years). The control subjects were recruited from the local community in Shanghai. They self-reported to be free from mental disorders, family history of mental illness and severe physical disorders. All patients and controls were ethnically Chinese Han. The study protocol and process were assessed and approved by the ethics committee at Shanghai Mental Health Center (IRB No. 2019-16R). We obtained informed consent from each participant.
Construction of protein-protein interaction network
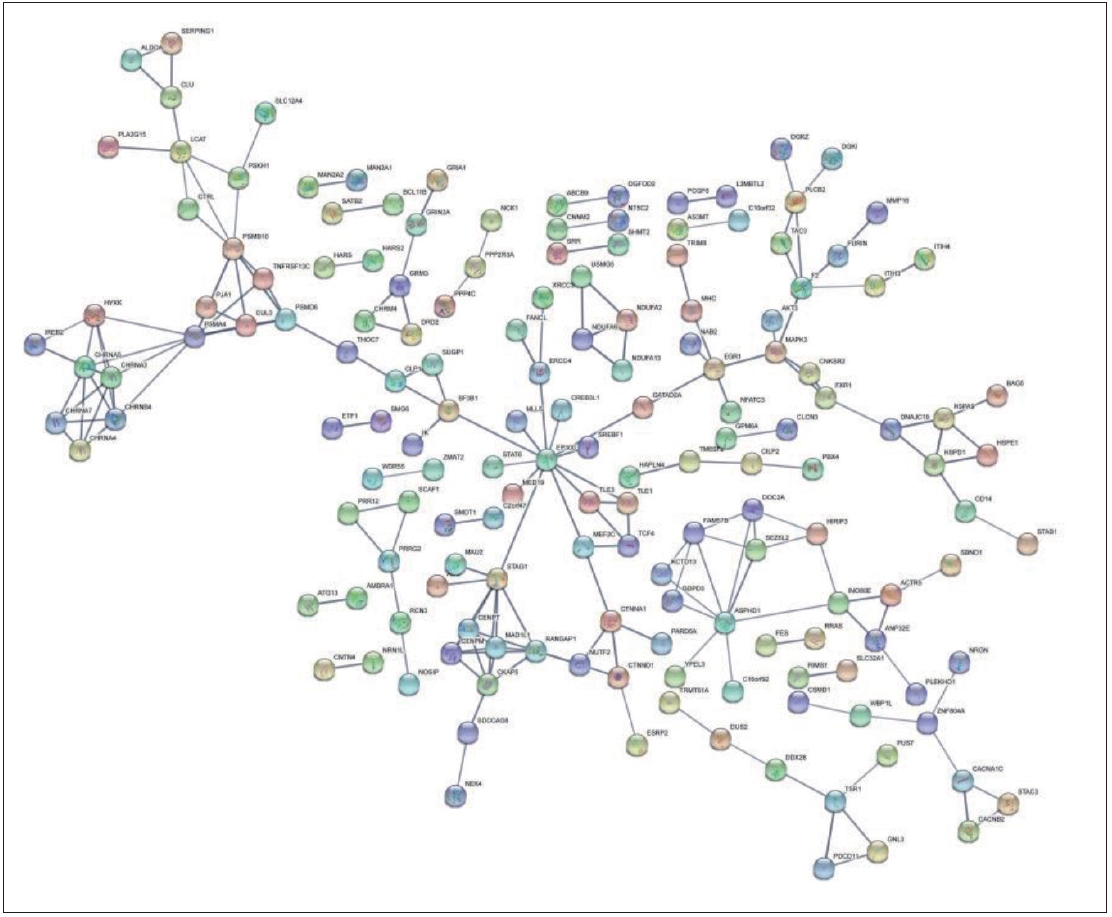
The Multiple proteins module of the CHRNA3, CHRNA4, CHRNA5, and CHRNA7 genes and 350 schizophrenia susceptibility genes identified in the Genome-wide Association Study (GWAS) of SZDB2.0 database (http://www.szdb.org) were uploaded to the String database (http://www.string-db.org/) [26,27]. A protein-protein interaction (PPI) network among the proteins encoded by these genes was constructed to determine the potential role of the four genes in the pathogenesis of schizophrenia. The String database is mainly used to obtain the PPI between gene-coding proteins by using previous data from different sources and can integrate the scores of different aspects of PPI to form a comprehensive score to represent the size of their interaction. In this study, the data of high confidence score of 0.7 was selected to construct the PPI network.
SNP selection and genotyping
A total of 5 SNPs within CHRNA3 (rs1317286), CHRNA4 (rs1044396), CHRNA7 (rs6494212), and CHRNA5 (rs16969968, rs684513) were selected based on the following criteria: 1) SNPs are capable of tagging more SNPs based on the linkage disequilibrium (LD) pattern of the respective genes according to the data from the HapMap CHB dataset (http://hapmap.ncbi.nlm.nih.gov/) and 1,000 Genomes (http://www.broadinstitute.org/mpg/snap/) and 2) all eligible SNPs should have a minor allele frequency (MAF) >5% according to the HapMap CHB and dbSNP datasets (http://www.ncbi.nlm.nih.gov/SNP/).
Genomic DNA was isolated using a Tiangen DNA isolation kit (Tiangen Biotech, Beijing, China). Genotyping of the SNPs was carried out using the TaqMan SNP Genotyping Assay (Applied Biosystems, Foster City, CA, USA) on ABI PRISM7900 sequence detection system instrument (Applied Biosystems) and SDS 2.0 software (Applied Biosystems). For quality control, all genotypes were determined without knowledge of case or control status in the genotyping process. 5% of the samples were repeated for genotyping and the corresponding results were 100% concordant.
Statistical analysis
Hardy-Weinberg equilibrium (HWE), allele and genotype frequency analyses and haplotype analysis were performed using SHEsis (http://shesisplus.bio-x.cn/SHEsis.html). SNPs with a p value less than 0.001 were considered as a departure from the HWE [28]. The interaction between the markers was analyzed using multifactor dimensionality reduction (MDR) software (version 3.0.2 (Computational Genetics Laboratory, University of Pennsylvania, State College, PA, USA). Power analysis was performed using the Quanto program (Version 1.2.3, https://bio.tools/QUANTO). All tests were two-tailed, and the significance level was set at 0.05.
RESULTS
The PPI network encoded by CHRNA3, CHRNA4, CHRNA5, CHRNA7 and schizophrenia susceptibility genes consists of 339 nodes and 191 edges. Each node represents a protein, and the edges represent interactions between proteins. As shown in the Figure 1, the proteins of CHRNA3, CHRNA4, CHRNA5, and CHRNA7 interact with the proteins encoded by the four schizophrenic-related susceptibility genes, which are iron-responsive element-binding protein 2 (IREB2), hydroxylysine kinase (HYKK9), proteasome subunit alpha type-4 (PSMA4), neuronal acetylcholine receptor subunit beta-4 (CHRNB4).
For all the SNPs investigated in the present study, there was no deviation from the Hardy-Weinberg equilibrium (p>0.05). There was no significant difference in either allele or genotype frequencies between the patients and controls in single marker analysis (Table 1). The haplotype structures of 4 SNPs (rs1317286, rs16969968, rs6494212, and rs684513) distributed on chromosome 15 were analyzed. The red square of the linkage disequilibrium (LD) block indicated a strong linkage between the marker rs16969968 and rs1317286 (DŌĆ▓=0.96), rs16969968 and rs684513 (DŌĆ▓=0.95), as shown in Figure 2. The haplotypes construct by markers 4 SNPs were not associated with schizophrenia (Table 2).
In interaction analysis, a significant association was revealed between models made up of rs1317286, rs1044396, rs6494212, and rs684513 and schizophrenia (p<0.05). Testing Balanced accuracy of the model is 0.4864, the result of Cross-Validation consistency is 10/10 (Table 3). Power analysis showed greater than 80% power (alpha=0.05) for schizophrenia samples with an OR=1.5, under the assumption of a log-additive model and disease prevalence of 1%. Thus, the likelihood of type II error appeared below.
DISCUSSION
This study does not demonstrate a significant genetic difference in 5 markers in nAChR genes between schizophrenia patients and healthy controls in the single marker analysis. The haplotypes constructed by these four markers of CHRNA3, CHRNA5, and CHRNA7 gene were not associated with schizophrenia either. However, analyzing the interaction of the SNPs by MDR, we found that the interaction between the four-SNP model, rs1317286, rs6494212, rs684513, and rs1044396 produced a significant risk effect for schizophrenia.
We selected four nicotinic receptor subunit genes in this study. CHRNA4 is located on chromosome 20 at position 20q13.2-q13.3. Some SNPs of CHRNA4 were found to be associated with many psychiatric disorders, such as schizophrenia, AlzheimerŌĆÖs disease (AD), attention-deficit/hyperactivity disorder (ADHD), ParkinsonŌĆÖs disease (PD), and nicotine dependence [29,30]. More specifically, a single nucleotide polymorphism in the CHRNA4 gene (rs1044396) appears to affect visual and auditory function, affecting speed and attention [31]. CHRNA3 is located on chromosome 15 at position 15q24. Recently, rs1051730 and rs1317286 of CHRNA3 were found to be associated with prepulse inhibition, which is the endophenotype of schizophrenia [23]. CHRNA5 is located on chromosome 15 at position 15q24. Association has been reported between schizophrenia and SNPs of the CHRNA5 [32].
The CHRNA7 located on chromosome 15q13-q14 was first genetically linked to the P50 auditory sensory processing deficit in schizophrenia [15]. Associations also have been reported between nicotine dependence and SNPs of the CHRNA7 (rsl909884, rs883473, rs2337980, rs6494212, rs904951, rs4779565, and rs10438287), and rs6494212 of CHRNA7 was reported to be associated with schizophrenia recently [33].
The PPI network found that the proteins of CHRNA3, CHRNA4, CHRNA5, and CHRNA7 interact with the proteins encoded by four schizophrenic-related susceptibility genes (REB2, HYKK9, PSMA4, and CHRNB4). To some extent, it suggested that nAChR protein might be involved in or affect the molecular pathway of schizophrenic-related susceptibility genes. The above findings suggested that the nAChR genes are possibly involved in the pathogenesis of schizophrenia. Therefore, we set out to study the association between schizophrenia and nAChR genes. But the present study revealed no association between schizophrenia and SNPs in the single-marker analysis.
Schizophrenia is a complex psychiatric disease is associated with numerous genes that individually confer small and incremental risk to the phenotype, and epistatic interactions can enhance the pathogenic effects of alleles [34,35]. In our study, the results of MDR revealed that polymorphisms in CHRNA3 (rs1317286), CHRNA7 (rs6494212), CHRNA5 (rs684513), and CHRNA4 (rs1044396) have significant gene-gene interactions in schizophrenia. These SNPs have been previously suggested to contribute to schizophrenia. susceptibility. These results demonstrate an accumulation of evidence for the role of an interaction between CHRNA3, CHRNA4, CHRNA5 and CHRNA7 may conferring susceptibility to schizophrenia.
There are several possible explanations for the lack of association between schizophrenia and the selected SNPs, noted as follows: Firstly, SNPs of the nAChR genes we selected are unlikely to play a substantial role in conferring susceptibility to schizophrenia in the Han Chinese population. But, associations of SNPs with schizophrenia or endophenotypes of schizophrenia have been reported across different ethnic groups. This could be explained by population differences where heterogeneous effects of the same polymorphism are caused by differences in genetic background. Secondly, schizophrenia is a complex disorder, in which multiple loci act jointly to confer susceptibility for disease. It is believed that schizophrenia involves multiple genetic and environmental causes. The genetic variation subjected to scrutiny in this study may provide only a limited contribution to schizophrenia. There is still a possibility that association occurs in subsets of schizophrenic patients. Further studies in a largely homogenous population are needed. Interestingly, when the SNPs were analyzed together, the interaction between four SNPs produced a significant risk effect for schizophrenia. Further research including family-based studies is needed to investigate the role of these genes in schizophrenia and the functions of the brain.
One limitation of this study is that we explored only one variant for CHRNA3, CHRNA4, and CHRNA7 and two variants for CHRNA5. Thus additional polymorphisms in these four subunit genes should be genotyped to confirm the interaction. Another limitation is that we did not analyze gene-environmental interaction in this study.
In conclusions, the results of the present study do not support polymorphisms selected as a major genetic susceptibility factor in schizophrenia in the single marker analysis. However, found that a four-SNP model had an important effect on schizophrenia by MDR analysis. Our findings may represent a significant genetic clue for the etiology of schizophrenia and thus further studies are required to uncover the exact role of genes in schizophrenia.